


тип хордовые. Общая характеристика и жизнедеятельность
Общая характеристика
Тип хордовых объединяет животных, весьма разнообразных по внешнему виду, образу жизни и условиям обитания. Представители хордовых встречаются во всех основных средах жизни: в воде, на поверхности суши, в толще почвы и, наконец, в воздухе. Географически они распространены по всему земному шару. Общее число видов современных хордовых равно примерно 40 тыс.
В тип хордовых входят бесчерепные (ланцетники), круглоротые (миноги и миксины), рыбы, земноводные, пресмыкающиеся, птицы и млекопитающие. К хордовым, как показали блестящие исследования А.О. Ковалевского, относится и своеобразная группа морских, а значительной мере сидячих животных – оболочников (аппендикулярии, асцидии, сальпы). Некоторые признаки сходства с хордовыми обнаруживает немногочисленная группа морских животных – кишечнодышащие, которых иногда также включают в тип хордовых.
Несмотря на исключительное разнообразие хордовых, все они обладают рядом общих черт строения и развития. Основные из них таковы:
1. У всех хордовых имеется осевой скелет, первоначально возникающий в виде спинной струны, или хорды. Хорда представляет собой упругий нечленистый тяж, эмбрионально развивающийся путем отшнуровывания его от спинной стенки зародышевой кишки. Таким образом, хорда имеет энтодермальное происхождение.
Последующая судьба хорды различна. Пожизненно она сохраняется только у низших хордовых (за исключением асцидий и сальи). Однако и в этом случае у большинства хорда в той или иной мере редуцируется в связи с развитием позвоночного столба. У высших хордовых она является эмбриональным органом и у взрослых животных в той или иной мере вытесняется позвонками, в связи с этим осевой скелет из сплошного, нечленистого становится сегментированным. Позвоночник, как и все другие скелетные образования (кроме хорды), имеет мезодермальное происхождение.
2. Над осевым скелетом располагается центральная нервная система, представленная полой трубкой. Полость нервной трубки носит название невроцеля. Трубчатое строение центральной нервной системы характерно практически для всех хордовых. Исключение составляют лишь взрослые оболочники.
Почти у всех хордовых передний отдел нервной трубки разрастается и образует головной мозг. Внутренняя полость сохраняется в этом случае в виде желудочков головного мозга.
Эмбрионально нервная трубка развивается из спинной части эктодермального зачатка.
3. Передний (глоточный) отдел пищеварительной трубки сообщается с наружной средой двумя рядами отверстий, получивших название жаберных щелей, так как у низших форм на их стенках располагаются жабры. Жаберные щели пожизненно сохраняются только у водных низших хордовых. У остальных они появляются лишь как эмбриональные образования, функционирующие на некоторых стадиях развития или не функционирующие вовсе.
Наряду с указанными тремя основными особенностями хордовых должны быть упомянуты следующие характерные черты их организации, которые, однако, кроме хордовых, имеются и у представителей некоторых других групп.
1. Хордовые, так же как и иглокожие, имеют вторичный рот. Он образуется путем прорыва стенки гаструлы на конце, противоположном гастропору. На месте же зарастающего гастропора формируется анальное отверстие.
2. Полость тела у хордовых вторичная (целом). Этот признак сближает хордовых с иглокожими и кольчатыми червями.
3. Метамерное расположение многих органов особенно четко выражено у зародышей и низших хордовых. У высших их представителей метамерия выражена слабо в связи с общим усложнением строения.
Наружной сегментации у хордовых нет.
4. Хордовым свойственна двусторонняя (билатеральная) симметрия тела. Как известно, этим признаком, кроме хордовых, обладают некоторые группы беспозвоночных животных.
Класс: млекопитающие
Общая характеристика
Млекопитающие – наиболее высокоорганизованный класс позвоночных животных. Основные прогрессивные черты млекопитающих следующие:
1) высокое развитие центральной нервной системы, в первую очередь серой коры полушарий большого мозга – центра высшей нервной деятельности. В связи с этим приспособительные реакции млекопитающих на условия среды весьма сложны и совершенны;
2) живорождение и выкармливание детенышей продуктом материнского организма – молоком, что позволяет млекопитающим размножаться при крайне разнообразных условиях жизни;
3) высокоразвитая способность к терморегуляции, обусловившая относительную температуру тела. Это вызвано, с одной стороны, регуляцией теплообразования (путем стимулирования окислительных процессов — так называемая химическая терморегуляция), с другой — регулированием отдачи тепла путем изменения характера кожного кровоснабжения и. силы испарения воды при дыхании и потоотделении (так называемая физическая терморегуляция.
Огромное значение в регулировании отдачи тепла имеет шерстный покров, а у некоторых и подкожный жировой слой.
Эти особенности, равно как и ряд других черт организации, обусловили возможность широкого распространения млекопитающих в самых разнообразных условиях. Географически они распространены почти повсеместно, за исключением Антарктиды. Еще более важно учесть, что млекопитающие заселяют самые разнообразные жизненные среды. Помимо многочисленных наземных видов, есть виды летающие, полуводные, водные и, наконец, заселяющие толщу почвы. Общее число видов современных млекопитающих равно примерно 4,5 тыс.
Морфологически млекопитающие характеризуются следующими признаками. Тело покрыто шерстью (исключения редки и имеют вторичный характер). Кожа богата железами. Особо должны быть отмечены млечные железы. Череп сочленяется с позвоночником двумя затылочными мыщелками. Нижняя челюсть состоит только из зубной кости. Квадратная и сочленовная кости превращаются в слуховые косточки и располагаются в полости среднего уха. Зубы дифференцированы на резцы, клыки и коренные: они сидят в альвеолах.. Локтевой сустав направлен назад, коленный – вперед, в отличие от низших наземных позвоночных, у которых оба эти сустава направлены вбок наружу (рис 1) Сердце четырехкамерное, сохраняется одна левая дуга аорты. Эритроциты безъядерны.
Строение млекопитающих
Кожный покров (рис. 1) у млекопитающих имеет более сложное строение, чем у других позвоночных. Сложно и разнообразно и его значение. Вся система кожных покровов играет огромную роль в терморегуляции млекопитающих Шерстный покров, а у водных видов (киты, тюлени) подкожный слой жира предохраняют тело от излишней потери тепла. Исключительно большую роль играет система кожных кровеносных сосудов. Диаметр их просветов регулируется нервнорефлекторным путем и может меняться в очень больших пределах. При расширении сосудов кожи теплоотдача резко увеличивается, при сужении, наоборот, сильно сокращается.
Большое значение для охлаждения организма имеет также испарение с поверхности кожи воды, выделенной потоковыми железами.
Благодаря описанным механизмам температура тела y многих млекопитающих относительно постоянна, и ее отличие от температуры внешней среды может быть равно примерно 100 0С. Так, у песца, обитающего зимой при температуре до —60 °С, температура тела равна примерно +39 °С. Следует, однако, иметь в виду, что постоянство температуры тела (гомойотермия) не является абсолютным признаком всех млекопитающих. В полной мере она характерна для плацентарных зверей, имеющих относительно крупные размеры.
У низших млекопитающих, у которых менее развит терморегуляционный механизм, и у мелких плацентарных зверей, имеющих невыгодное для сохранения тепла соотношение между объемом тела и его поверхностью, температура тела меняется в значительных пределах в зависимости от температуры внешней среды (рис. 3). Так у сумчатой крысы температура тела меняется в пределах +37,8... +29,3 °С, у наиболее примитивных насекомоядных (тенреков) 4-34... 4- 13 °С, у одного из видов броненосцев 4-40... +27 ЭС, у обыкновенной полевки +37... +32 °С.
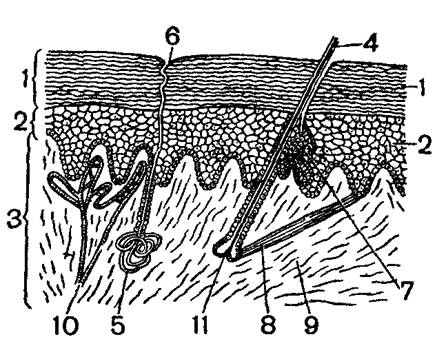
Рис. 2. Строение кожи млекопитающего (сильное увеличение)

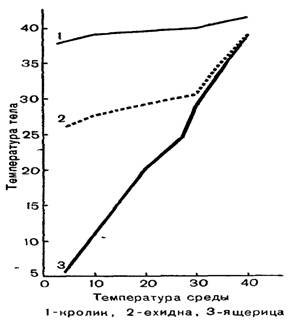
Рис.3. Кривые зависимости температуры тела различных животных от температуры внешней среды
Как и у других позвоночных, кожа млекопитающих состоит из двух слоев: наружного – эпидермиса и внутреннего – кутиса, или собственно кожи. Эпидермис в свою очередь состоит из двух слоев. Глубокий слой, представленный живыми цилиндрическими или кубическими клетками, известен под названием мальпигиева или росткового слоя. Ближе к поверхности клетки более плоские, в них появляются включения кератогиалина, который, постепенно заполняя полость клетки, ведет к ее роговому перерождению и отмиранию. Поверхностно расположенные клетки окончательно ороговевают и постепенно снашиваются в виде мелкой «перхоти» или целыми лоскутами (так, например, бывает у тюленей). Снашивание рогового слоя эпидермиса выполняется постоянным нарастанием его за счет деления клеток мальпигиева слоя.
Эпидермис дает начало многим производством кожи, основные из которых – волосы, когти, копыта, рога (кроме оленьих), чешуи, различные железы. Образования эти описаны ниже.
Собственно кожа, или кутис, у млекопитающих развита очень сильно. Она состоит преимущественно из волокнистой соединительной ткани, сплетения волокон которой образуют сложный узор. Нижняя часть кутиса состоит из очень рыхлой волокнистой ткани, в которой откладывается жир. Слой этот носит название подкожной жировой клетчатки. Наибольшего развития он достигает у водных зверей – китов, тюленей, у которых он в связи с полной (у китов) или частичной (у тюленей) редукцией волосяного покрова и физическими особенностями водной среды выполняет термоизоляционную роль. У некоторых наземных зверей также бывают большие подкожные жировые отложения. Особенно сильно они развиты у видов, впадающих на зиму в спячку (суслики, сурки, барсуки и др.). Для них жир во время спячки служит основных энергетическим материалом.
Толщина кожи существенно различна в разных видов. Как правило, у видов холодных стран, имеющих пышный волосяной покров, она толще. Очень тонкая и непрочная кожа характерна для зайцев, к тому же она бедна кровеносными сосудами. Это имеет известное приспособительное значение, выражающееся в своеобразной автономии. Хищник, схвативший зайца за кожу, легко вырывает из нее кусок, упуская самого зверька. Образовавшаяся рана почти не кровоточит и быстро заживает. Своеобразная кожная хвостовая автономия наблюдается у некоторых мышей, сонь, тушканчиков. Кожный хвостовой футляр у них легко обрывается и соскальзывает с хвостовых позвонков, что дает возможность схваченному за хвост зверьку уйти от врага.
Волосяной покров для млекопитающих столь же характерен, как покров из перьев для птиц или чешуйчатый покров для рептилий. Лишь немногие виды вторично полностью или частично утратили волосяной покров. Так, вовсе не имеют волос дельфины, у китов есть только задатки волос на губах. У ластоногих волосяной покров редуцированный, особенно это заметно у моржей, в наименьшей мере – у ушатых тюленей (например, у котика), которые более, чем другие виды ластоногих, связанны с сушей.
Строение волоса видно в схеме на рисунке 2. В нем можно различить ствол – часть, выступающую над кожей, и корень – часть, сидящую в коже. Ствол состоит из сердцевины, коркового слоя и кожицы. Сердцевина представляет собой пористую ткань, между клетками которой находится воздух; именно эта часть волоса сообщает ему малую теплопроводность. Корковый слой, наоборот, очень плотный и придает волосу прочность. Тонкая наружная кожица защищает волос от механических и химических повреждений. Корень волоса в своей верхней части имеет цилиндрическую форму и является прямым продолжением ствола. В нижней части корень расширяется прямым продолжением ствола. В нижней части корень расширяется и заканчивается колбообразным вздутием – луковицей волоса, которая, как колпачок, охватывает вырост кутиса – волосяной сосочек. Кровеносные сосуды, входящие в этот сосочек, обеспечивают жизнедеятельность клеток луковицы волоса. Формирование и нарастание волос идет за счет размножения и видоизменения клеток луковицы. Ствол волоса является уже мертвым роговым образованием, неспособным к росту и изменению формы.
Погруженный в кожу корень волоса сидит в волосяной сумке, стенки которой состоят из наружного слоя, или волосяного мешка, и внутреннего слоя, или волосяного влагалища. В воронку волосяного мешка открываются протоки сальных желез, секрет которых смазывает волос и сообщает ему большую прочность и водоупорность. К нижней части волосяного мешка прикрепляются мышечные волокна, сокращения которых вызывают движение мешка и сидящего в нем волоса. Это движение и обуславливает ощетинивание зверя.
Обычно волосы сидят в коже не перпендикулярно ее поверхности, а более или менее прилегают к ней. Такой наклон волос выражен не у всех видов одинаково. Наименее заметен он у подземных зверей, например у крота.
Волосяной покров состоит из различных типов волос. Основные из них – это пуховые волосы, или пух, остевые волосы, или ость, чувствующие волосы, или вибриссы. У большинства видов основу шерстного покрова составляет густой низкий пух, или подшерсток. Между пуховыми волосами сидят более длинные, толстые и жесткие остевые волосы. У подземных зверей, например у крота, слепыша, меховой покров почти всегда лишен остевых волос. Наоборот, у взрослых оленей, кабанов и тюленей редуцирован подшерсток и покров состоит главным образом из ости. Отметим, что у молодых особей этих зверей подшерсток развит хорошо.
Волосяной покров периодически меняется. Смена волос, или линька, у некоторых видов бывает два раза в году: весной и осенью: таковы белка, лисица, песец, крот. Другие виды линяют только раз в год; весной у них выпадает старый мех, летом развивается новый, который окончательно созревает только к осени. Таковы, например, суслики.
Густота и высота волосяного покрова у северных видов существенно меняются по сезонам. Так, у белки на 1 см2 на огузке летом бывает в среднем 4200 волос, зимой 8100, то же у зайца-беляка – 8000 и 14700. Длина волос в миллиметрах на огузке такова: у белки летом пух – 9,4 ость – 17,4, зимой – 16,8 и 25,9; у зайца беляка летом пух – 12,3, ость – 26,4, зимой 21,0 и 33,4. У тропических зверей таких резких изменений не бывает в силу малой разницы в температурных условиях зимой и летом.
Особую категорию волос представляют вибриссы. Это очень длинные жесткие волосы, выполняющие осязательную функцию, сидят чаще на голове (так называемые усы), на нижней части шеи, на груди, а у некоторых лазающих древесных форм (например, у белки) и на брюхе. В основании волосяного мешка и в его стенках располагаются нервные рецепторы, воспринимающие соприкосновение стержня вибриссы с посторонними предметами.
Видоизменениями волос являются щетины и иглы.
Прочие роговые производные эпидермиса представлены чешуями, ногтями, когтями, копытами, полыми рогами, роговым клювом. Чешуя зверей по своему развитию и строению вполне сходна с одноименным образованием рептилий. Наиболее сильно развита чешуя у ящеров и панголинов, у которых она покрывает все тело. У очень многих мышевидных чешуйки имеются на лапках. Наконец, наличие чешуи на хвосте характерно для многих сумчатых, грызунов и насекомоядных.
Концевые фаланги пальцев подавляющего большинства зверей несут роговые придатки в виде ногтей, когтей или копыт. Наличие того или иного из этих образований и строение их находятся в прямой связи с условиями существования и образом жизни зверей (рис.4). Так, у лазающих зверей пальцы имеют острые загнутые когти; у видов, роющих в земле норы, когти обычно несколько упрощены и расширены. Быстро бегающие крупные млекопитающие имеют копыта, при этом у лесных видов (например, у оленей), часто ходящих по болотам, копыта более широкие и плоские. У степных (антилопы) и особенно у горных видов (козлы, бараны) копыта маленькие, узкие; площадь опоры у них значительно меньше, чем у лесных копытных, ходящих зачастую по размягченному грунту или по снегу. Так, нагрузка на 1 см2 подошвы у центрально-азиатского горного козла равна в среднем 850 г., у лося – 500 г, у северного оленя – 140 г.
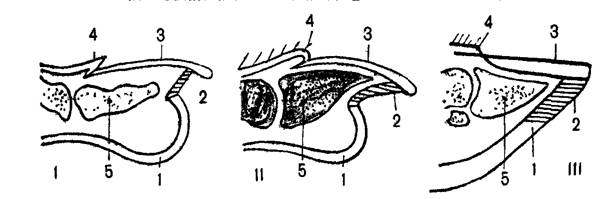
Рис. 4. Продольный разрез через конечные фаланги пальцев обязьяны (1), хищника (II), копытного (III):
Роговыми образованиями являются также рога быков, антилоп, козлов и баранов. Они развиваются из эпидермиса и сидят на костных стержнях, представляющих самостоятельные кости, сросшиеся с лобными костями. Рога оленей имеют иную природу. Они развиваются из кутиса и состоят из костного вещества.
Кожные железы у млекопитающих, в отличие от птиц и рептилий, весьма многочисленны и разнообразны по строению и функции. Основные типы желез следующие: потоковые, сальные, пахучие, млечные.
Потовые железы трубчатые, глубинные части их имеют вид клубка. Они открываются непосредственно из поверхности кожи или в волосяную сумку. Продуктом выделения этих желез является пот, состоящий в основном из воды, в которой растворены мочевина и соли. Эти продукты не вырабатываются клетками желез, а поступают в них из кровеносных сосудов. Функция потовых желез заключается в охлаждении тела путем испарения выделяемой ими на поверхности кожи воды и в выделении продуктов распада. Следовательно, эти железы выполняют терморегуляционную функцию. Потовые железы есть у большинства млекопитающих, но развиты они не у всех одинаково. Так, их очень мало у собак и кошек; у многих грызунов они имеются только на лапах, в паховой области и на губах. Совершенно отсутствуют потовые железы у китообразных, ящеров и некоторых других.
В развитости потовых желез можно подметить и закономерности географического и экологического планов. Так, среднее количество этих желез на 1 см2 у разводимого во влажных тропиках зебу равно 1700, а у крупного рогатого скота, разводимого в Англии (шортхорн), - всего 1060. Эта же особенность прослеживается при сравнении видов, в разной мере приспособленных к засушливым условиям. В качестве показателя приводим величину испарения, выраженную в миллиграммах в минуту на 100 см2 поверхности кожи. При температуре +37 0С у осла эта величина равнялась 17 мг/мин, у верблюда – только 3; при температуре +45 0С у осла – 35, у верблюда – 15; наконец, при температуре +50 0С у осла – 45, у верблюда – 25 (Шмидт-Нильсен, 1972).
Секрет кожных желез, как и другие пахнущие выделения (например, полового и пищеварительного трактов, моча, секрет специализированных желез), служат наиболее важным средством внутривидового общения – химической сигнализации млекопитающих. Особое значение этого типа сигнализации определяется дальностью ее действия и длительностью сохранения сигнала. У животных, имеющих определенные участки обитания, особи, пары, семьи метят участок запаховыми метками, которые они оставляют на приметных предметах: кочках, камнях, пнях, отдельных деревьях или просто на поверхности земли.
Сальные железы имеют гвоздевидное строение и открываются почти всегда в воронку волосяной сумки. Жирный секрет этих желез смазывает волос и поверхностный слой эпидермиса кожи, предохраняя их от смачивания и снашивания.
Пахучие железы представляют видоизменение потовых или сальных желез, а иногда комбинацию тех и других. Из числа их укажем на анальные железы куньих, секрет которых имеет очень резкий запах.
Запаховые метки оставляют родители на детенышах, в гнезде и на следах движения за пределами гнезда или места нахождения детеныша, если гнездо не сооружается. Именно благодаря химической сигнализации олени, тюлени и такие норники, как лисицы, песцы, соболи, куницы, полевки, мыши, находят своих, а не чужих детенышей.
В целом запаховая сигнализация имеет решающее значение для выработки форм поведения млекопитающих.
Особенно сильно развиты пахучие железы у американских скунсов, или вонючек (Mephitis), способных выпрыскивать большие порции выделений на значительное расстояние. Мускусные железы имеются у .кабарги, выхухоли, бобра, ондатры; значение этих желез не вполне ясно, но, судя по тому, что наибольшее развитие они получают в период гона, деятельность их, видимо, связана с размножением; возможно, они стимулируют половое возбуждение.
Млечные железы представляют своеобразное видоизменение простых трубчатых потовых желез. В простейшем случае — у австралийских однопроходных — они сохраняют трубчатое строение и открываются в сумки волос, расположенных группами на небольшом участке брюшной поверхности — так называемом железистом поле. У ехидны железистое поле находится в особой сумке, развивающейся в период размножения и служащей для вынашивания яйца, а потом и детеныша. У утконоса железистое поле располагается непосредственно на брюхе. Сосков у однопроходных нет и детеныши слизывают молоко с волос, куда оно поступает из волосяных сумок. У сумчатых и плацентарных млечные железы имеют гроздевидное строение и протоки их открываются на сосках . Расположение желез и сосков бывает различным. У лазающих по деревьям обезьян в висящих во время кормленая летучих мышей имеется только пара qockob на груди; у бегающих копытных соски располагаются только в паховой области. У насекомоядных и хищных соски тянутся двумя рядами по всей нижней поверхности туловища. Число сосков стоит в прямой связи с плодовитостью вида и в некоторой степени соответствует числу одновременно рождающихся детенышей. Минимальное число сосков (2) характерно для обезьян, овец, коз, слонов и некоторых других; максимальное число сосков (10 — 24) свойственно мышевидным грызунам, насекомоядным, некоторым сумчатым.
Мышечная система млекопитающих очень дифференцированна и отличается большим числом разнообразно расположенных мускулов. Характерно наличие куполообразной мышцы — диафрагмы, ограничивающей брюшную полость от грудной. В основном ее роль заключается в изменении объема грудной полости, что связано с актом дыхания. Значительное развитие получает подкожная, мускулатурная, приводящая в движение те или иные участки кожи. У ежей и ящеров она обуславливает возможность свертывания тела в клубок. Поднятие игл у ежей и дикобразов, «ощетинивание» зверей и движение чувствующих волос – вибрисс – также вызываются действием мускулатуры. На лице она представлена мимической мускулатурой, особенно развитой у приматов.
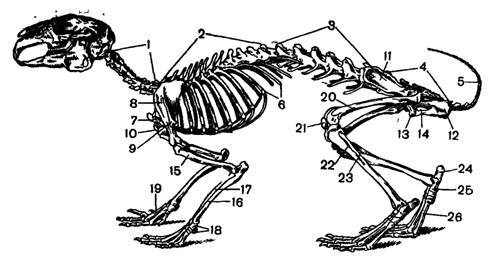
Рис. 5. Скелет кролика

Скелет. (рис. 5). Характерными чертами в строении позвоночного столба млекопитающих являются плоские сочленовные поверхности позвонков (платицельные позвонки), между которыми располагаются хрящевые диски (мениски), четко выраженная расчлененность позвоночника на отделы (шейный, грудной, поясничный, крестцовый, хвостовой) и постоянное число швейных позвонков. Отклонения от указанных признаков редки и носят вторичный характер.
В шейном отделе характерно наличие хорошо выраженных атланта и эпистрофея – видоизмененных двух первых позвонков, что типично вообще для амниот. Шейных позвонков 7. Исключение представляют только ламантин, у которого 6 шейных позвонков, и виды ленивцев, имеющих от 6 до 10 позвонков. Таким образом, в отличие от птиц, у млекопитающих длина шеи определяется не числом шейных позвонков. А длиной их тела. Длина шейного отдела варьирует очень значительно. Наиболее сильно он развит у копытных, для которых подвижность головы очень важна при добыче пищи. Хорошо развита шея у хищников. Наоборот, у грызунов-норников и особенно у землероев шейный отдел короток и подвижность головы у них малая.
Грудной отдел состоит чаще из 12 – 15 позвонков; у одного из броненосцев и клюворылого кита их 9, а у ленивцев рода Choloepus – 24. К передним грудным позвонкам обычно к семи и причленяются ребра, соединенные с грудиной (истинные ребра). Остальные грудные позвонки несут ребра, не доходящие до грудины (ложные ребра). Грудина представляет сегментированную костную пластинку, заканчивающуюся удлиненным хрящом – мечевидным отростком. Расширенный передний сегмент носит название рукоятки грудины. У летучих мышей и у зверей с хорошо развитыми для рытья передними конечностями грудина теряет ясно выраженную сегментацию и несет киль, служащий, как и у птиц, для прикрепления грудных мышц.
В поясничном отделе число позвонков варьирует от 2 до 9. Позвонки эти несут рудиментарные ребра.
Крестцовый отдел состоит чаще из четырех сросшихся позвонков. При этом только первые два позвонка являются истинно крестцовыми, а остальные – приросшими к крестцу хвостовыми позвонками. У жищных число крестцовых позвонков равно трем. А у утконоса, как и рептилий – двум. Число хвостовых позвонков подвержено наибольшей изменчивости. Так, у гиббона из 3, а у длиннохвостого ящера – 49.
Общая подвижность позвоночника у разных видов зверей различна. Наиболее сильно она развита у мелких зверьков, которые при движении часто выгибают спину дугой. Наоборот, у крупных копытных все отделы позвоночника (кроме шейного и хвостового) перемещаются незначительно, и при беге их работают только конечности.
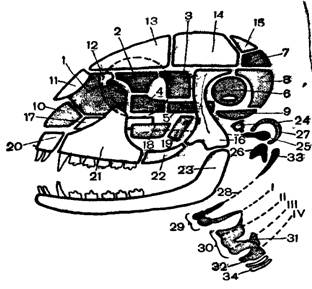
Рис. 6. Схема строения черепа млекопитающих

Череп млекопитающих (рис. 6) характерен относительно более крупной мозговой коробкой, что связано с большими размерами головного мозга. У молодых зверей мозговая коробка по сравнению с лицевой частью обычно развита относительно сильнее, чем у взрослых. Число отдельных костей в черепе млекопитающих меньше, чем у нижестоящих групп позвоночных. Это обуславливается срастанием ряда костей между собой, что особенно характерно для мозговой коробки. Так, срастаются основная, боковые и верхняя затылочная кости; сращение ушных костей приводит к образованию единой каменистой кости. Крылоклиновидная кость срастается с основной клиновидной костью, а глазоклиновидная кость срастается с передней клиновидной костью. Бывают случаи образования и более сложных комплексов, например височная и основная кости человека. Швы между костными комплексами срастаются сравнительно поздно, особенно в области мозговой коробки, что дает возможность увеличения объема головного мозга по мере роста животного.
Затылочная область сформирована единой, как указано, затылочной костью, имеющей два мыщелка для сочленения с атлантом. Крыша черепа образована парными теменными, лобными и носовыми костями и непарной межтеменной костью. Бока черепной коробки образованы чешуйчатыми костями, от которых наружу и вперед отходят скуловые отростки. Последние соединяются со скуловой костью, которая в свою очередь спереди сочленена со скуловым отростком верхнечелюстной кости. В итоге образуется весьма характерная для млекопитающих скуловая дуга.
Дно мозговой части черепа образовано основной и переднеклиновидной костями, а дно висцеральной части – крыловидными, небными и верхнечелюстными костями. На дне черепа, в области слуховой капсулы, располагается свойственная только млекопитающим барабанная кость. Слуховые капсулы окостеневают, как уже указано, несколькими центрами, но в конечном счете формируется только одна парная каменная кость.
Верхние челюсти состоят из парных межчелюстных и верхнечелюстных костей. Характерно развитие вторичного костного неба, образованного небными отростками межчелюстных и верхнечелюстных костей и небными костями. В связи с образованием вторичного костного неба хоаны открываются не между верхнечелюстными костями, как у других наземных позвоночных (кроме крокодилов и черепах), а позади небных костей. Такое строение неба предотвращает закупорку хоан (т.е. перерыв дыхания) в то время, как пищевой комок задерживается в ротовой полости для его пережевывания.
Нижняя челюсть представлена только парными зубными костями, которые причленяются непосредственно к чешуйчатым костям. Сочленовная кость превращается в слуховую косточку – наковальню. Обе эти косточки, равно как и третья слуховая косточка – стремя (гомолог гиомандибуляре), лежат в полости среднего уха. Наружная стенка последней, а также часть наружного слухового прохода окружены упомянутой выше барабанной костью, видимо гомологичной угловой кости – нижней челюсти прочих позвоночных. Таким образом, у млекопитающих наблюдается дальнейшее превращение части висцерального аппарата в слуховой аппарат среднего и наружного уха.
Плечевой пояс млекопитающих сравнительно прост. Основой его является лопатка, к которой прирастает рудиментарный коракоид. Только у однопроходных коракоид существует в виде самостоятельной кости. Ключица имеется у млекопитающих, передние конечности которых совершают разнообразные сложные движения и у которых наличие ключицы обеспечивает более прочное причленение плечевой кости и упрочнение всего плечевого пояса. Таковы, например, обезьяны. Наоборот, у видов, которые перемещают передние конечности только или преимущественно в плоскости, параллельной главной оси тела, ключицы рудиментарны или отсутствуют. Таковы копытные.
Тазовый пояс состоит из трех типичных для наземных позвоночных парных костей: подвздошных, седалищных и лобковых. У многих видов эти кости срастаются в одну безымянную кость.
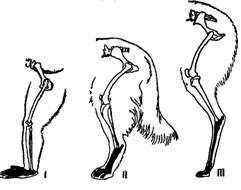
Рис.7. Задние конечности пальце- и стопоходящих млекопитающих.
Элементы стопы зачернены.
I – обезьяна павиан, II – собака, III – лама.
Скелет парных конечностей сохраняет все основные черты строения типичной пятипалой конечности. Однако в связи с разнообразием условий существования и характером использования конечностей детали их строения весьма различны (рис.7) У наземных форм значительно удлинены проксимальные отделы. У водных зверей, наоборот, эти отделы укорочены, а дистальные отделы — пясть, плюсна и особенно фаланги пальцев — сильно удлинены. Конечности в этом случае приращены в ласты, перемещающиеся относительно тела в основном как единое целое. Перемещение же отделов конечностей относительно друг друга развито сравнительно слабо. У летучих мышей только первый палец передних конечностей развит нормально, остальные пальцы очень сильно удлинены; между ними расположена кожистая перепонка, образующая основную часть поверхности крыла. У быстро бегающих зверей предплюсна, плюсна, запястье и пясть располагаются более или менее отвесно, и животные эти опираются только на пальцы. Таковы, например, собаки. У наиболее совершенных бегунов – копытных – сокращается число пальцев. Первый палец атрофируется, и животные ступают либо на одинаково развитые третий и четвертый пальцы, между которыми проходит ось конечности (парнокопытные), либо преимущественное развитие получает один третий палец, через который и проходит ось конечностей (непарнокопытные).
Укажем в этой связи максимальную скорость передвижения некоторых млекопитающих (в км/ч): короткохвостая землеройка – 4, красная полевка – 7, лесная мышь – 10, красная белка – 15, дикий кролик – 32-40, заяц – 55-72, красная лисица – 72, лев – 50, гепард – 105-112, верблюд – 15-16, африканский слон – 24-40, газель Гранта – 40-50.
Органы пищеварения характеризуются большой сложностью, которая выражается в общем удлинении пищеварительного тракта, в большей, чем у других позвоночных, его дифференцировке и в большей развитости пищеварительных желез.
Пищеварительный тракт начинается предротовой полостью или преддверием рта, расположенным между свойственными только млекопитающим мясистыми губами, щеками и челюстями. У ряда видов преддверие, расширяясь, образует большие защечные мешки. Так бывает у хомяков, бурундуков, обезьян. Мясистые губы служат для схватывания пищи, а преддверие рта – для временного ее резервирования. Так, хомяки и бурундуки переносят в защечных мешках запасы пищи в свои норы. Мясистых губ нет у однопроходных и китообразных.
За челюстями лежит ротовая полость, в которой пища подвергается механическому измельчению и химическому воздействию. У зверей имеются четыре пары слюнных желез, секрет которых содержит фермент птиалин, превращающий крахмал в декстрин и мальтозу. Развитость слюнных желез находится в известной зависимости от характера питания. У китообразных они практически не развиты; у жвачных, наоборот, они получили исключительно сильное развитие. Так, корова в сутки выделяет около 56 л слюны, которая имеет огромное значение для смачивания грубой пищи и для наполнения жидкой средой полостей желудка, где происходит бактериальное расщепление клетчатки пищевой массы.
Секрет щечных желез летучих мышей, наносимый на летательные перепонки, сохраняет их эластичность и предохраняет от высыхания. Слюна вампиров, питающихся кровью, обладает антикоагулирующими свойствами, т.е. предохраняет кровь от свертывания. Слюна некоторых землероек ядовита, секрет их подчелюстной железы вызывает смерть мыши менее через 1 мин после инъекции. Токсичность слюнных желез примитивных млекопитающих рассматривается как отражение их филогенетической связи с рептилиями.
Млекопитающие гетеродонтны, т.е. их зубы дифференцированы на резцы, клыки, предкоренные, или ложные коренные, и коренные. Число зубов, их форма и функция существенно различаются у разных групп зверей. Так, например мало специализированные насекомоядные обладают большим числом сравнительно слабо дифференцированных зубов. Для грызунов и зайцеобразных характерно сильное развитие одной пары резцов, отсутствие клыков и плоская жевательная поверхность коренных зубов. Такое строение зубной системы связано с характером питания: резцами они сгрызают или обгрызают растительность, а коренными зубами перетирают пищу, как жерновами. Хищные характеризуются сильно развитыми клыками, служащими для схватывания, а часто и для умерщвления добычи. Коренные зубы хищных имеют режущие вершины и плоские жевательные выступы. Задний ложнокоренной зуб верхней челюсти и первый истиннокоренной нижней челюсти у хищных обычно выделяются своей величиной; они носят название хищных зубов.
Общее число зубов и их распределение по группам для видов зверей вполне определено и постоянно и служит важным систематическим признаком.
Зубы сидят в ячейках челюстных костей, т.е. они текодонтные, и у большинства видов зверей меняются один раз в жизни (зубная система дифиодонтна).
Между ветвями нижней челюсти помещается мускулистый язык, служащий отчасти для схватывания пищи (бычьи, муравьеды, ящеры) и для лакания воды, отчасти для переворачивания пищи в полости рта во время ее пережевывания.
Позади ротовой области располагается глотка, в верхнюю часть которой открываются внутренние ноздри и евстахиевы трубы. На нижней поверхности глотки расположена щель, ведущая в гортань.
Пищевод хорошо выражен. Мускулатура его чаще гладкая, но у некоторых, например у жвачных, сюда проникает из глоточной области поперечнополосатая мускулатура. Эта особенность обеспечивает произвольное сокращение пищевода при отрыгивании пищи.
Желудок явственно обособлен от других отделов пищеварительного тракта и снабжен многочисленными железами. Объем желудка и его внутреннее строение различны у разных видов, что связано с характером пищи. Наиболее просто устроен желудок у однопроходных, у которых он имеет вид простого мешка. У большинства желудок разлен на большее или меньшее число отделов.
Усложнение желудка связано со специализацией питания, например поглощением огромной массы грубых кормов (жвачные), или недоразвитостью ротового пережевывания пищи (некоторые виды, питающиеся насекомыми). У некоторых южноамериканских муравьедов в выходной части желудка дифференцируется отдел с настолько твердыми складками, что они выполняют фун