


Энергетический метаболизм микроорганизмов
1. Общие понятия об обмене веществ и энергии
2. Конструктивный метаболизм
3. Потребность прокариот в питательных веществах
3.1 Источники углерода
3.2 Азот
3.3 Потребности в источниках серы и фосфора
3.4 Необходимость ионов металлов
3.5 Потребность в факторах роста
4. Типы метаболизма микроорганизмов
5. Энергетический метаболизм фототрофов
6. Энергетический метаболизм хемотрофов, использующих процессы брожения
7. Энергетический метаболизм хемоорганотрофов, использующих процесс дыхания
8. Энергетический метаболизм хемолитоавтотрофов
Заключение
Список литературы
Данная курсовая работа содержит основные сведенья о конструктивном и энергетическом метаболизме бактерий. Работа выполнена на 37 листах. Содержит 5 рисунков и 1 таблицу.
Совокупность процессов превращения материи в живом организме, сопровождающихся постоянным ее обновлением, называется обменном веществ или метаболизмом.
Важнейшими свойствами живых организмов являются способность, к самовоспроизведению и теснейшая взаимосвязь их с окружающей средой. Любой организм может существовать только при условии постоянного притока питательных веществ из внешней среды и выделения в нее продуктов жизнедеятельности.
Питательные вещества, поглощаемые клеткой, в результате сложных биохимических реакций превращаются в специфические клеточные компоненты. Совокупность биохимических процессов поглощения, усвоения питательных веществ и создания за их счет структурных элементов клетки называется конструктивным обменом или анаболизмом. Конструктивные процессы идут с поглощением энергии. Энергию, необходимую для процессов биосинтеза других клеточных функций, таких, как движение, осморегуляция и т. д., клетка получает за счет потока окислительных реакций, совокупность которых представляет собой энергетический обмен, или катаболизм (рис. 1).
1. Общие понятия об обмене веществ и энергии
Все живые организмы могут использовать только химически связанную энергию. Каждое вещество обладает определенным запасом потенциальной энергии. Главные материальные носители ее химические связи, разрыв или преобразование которых приводит к освобождению энергии.
Энергетический уровень химических связей неодинаков. Для одних он имеет величину порядка 8—10 кДж. Такие связи называют нормальными. В других связях заключена значительно большая энергия - 25—40 кДж. Это так называемые макроэргические связи. Почти все известные соединения, обладающие такими связями, включают атомы фосфора и серы, участвующие в образовании этих связей.
Важнейшую роль в жизнедеятельности клетки играет аденозинтрифосфорная кислота (АТФ). В состав ее молекулы входят аденин, рибоза и три остатка фосфорной кислоты: (Приложения Рис 2)
АТФ занимает центральное место в энергетическом обмене клетки. Макроэргические связи в молекуле АТФ очень непрочны. Гидролиз этих связей приводит к освобождению значительного количества свободной энергии:
АТФ + Н20→АДФ + Н3Р04- 30,56 кДж
Гидролиз протекает с участием специфических ферментов, обеспечивая энергией биохимические процессы, идущие с поглощением энергии. В этом случае АТФ играет роль поставщика энергии. Имея малый размер, молекула АТФ и диффундирует в различные участки клетки. Запас АТФ в клетках непрерывно возобновляется за счет реакций присоединения остатка фосфорной кислоты к молекуле аденозиндифосфорной кислоты (АДФ):
АДФ + Н3Р04 → АТФ + Н20
Синтез АТФ, как и гидролиз, идет при участии ферментов но сопровождается поглощением энергии, способы получения которой у микроорганизмов хотя и разнообразны, но могут быть сведены к двум типам:
1) использование энергии света;
2) использование энергии химических реакций.
При этом тот и другой виды энергии трансформируются в энергию химических связей АТФ. Таким образом, АТФ выполняет в клетке роль трансформатора.
Анаболизм и катаболизм неразрывно связаны, составляя единое целое, поскольку продукты энергетического обмена (АТФ и некоторые низкомолекулярные соединения) непосредственно используются в конструктивном обмене клетки (рис. 6.1).
В клетках микроорганизмов соотношение между энергетическими и конструктивными процессами зависит от ряда конкретных условий, в частности от характера питательных веществ. Тем не менее по объему катаболические реакции обычно превосходят биосинтетические процессы. Взаимосвязь и сопряженность этих двух видов метаболизма проявляется прежде всего в том, что суммарный объем конструктивных процессов полностью зависит от количества доступной энергии, получаемой в ходе энергетического обмена.
Конструктивный метаболизм направлен на синтез четырех основных типов биополимеров: белков, нуклеиновых кислот, полисахаридов и липидов.
Ниже показана обобщенная условная схема биосинтеза сложных органических соединений, где выделены следующие основные этапы: образование из простейших неорганических веществ органических предшественников (I), из которых на следующем этапе синтезируются «строительные блоки» (II). В дальнейшем строительные блоки, связываясь друг с другом ковалентными связями, образуют биополимеры (III): Приложения (рис. № 3)
Представленная схема биосинтетических процессов не отражает всей сложности превращения низкомолекулярных предшественников в строительные блоки с большой молекулярной массой. На самом деле синтез протекает как серия последовательных реакций с образованием разнообразных промежуточных продуктов метаболизма. Кроме того, уровни развития биосинтетических способностей микроорганизмов очень различны. У одних микробов конструктивный метаболизм включает все показанные на схеме этапы, у других ограничен вторым и третьим или только третьим этапом. Именно поэтому микроорганизмы резко отличаются друг от друга по своим пищевым потребностям. Однако элементный состав пищи одинаков для всех живых организмов и должен включать все компоненты, входящие в клеточное вещество: углерод, азот, водород, кислород и др.
В зависимости от используемых в конструктивном обмене источников углерода микроорганизмы делятся на две группы: автотрофы и гетеротрофы.
Автотрофы (от греч. «autos» — сам, «trophe» — пища) в качестве единственного источника углерода используют диоксид углерода и из этого простого неорганического соединения-предшественника синтезируют все необходимые биополимеры. Способность к биосинтезу у автотрофов самая высокая.
Гетеротрофы (от греч. «heteros» — другой) нуждаются в органических источниках углерода. Их пищевые потребности чрезвычайно разнообразны. Одни из них питаются продуктами жизнедеятельности других организмов или используют отмершие растительные и животные ткани. Такие микроорганизмы называются сапрофитами (от греч. «sapros» — гнилой и «phyton» — растение). Число органических соединений, используемых ими в качестве источников углерода, чрезвычайно велико — это углеводы, спирты, органические кислоты, аминокислоты и т. д. Практически любое природное соединение может быть использовано тем или иным видом микроорганизмов в качестве источника питания или энергии.
Вторую группу гетеротрофных организмов составляют паразиты, развивающиеся в живых клетках. Паразиты, нарушая равновесие биохимических процессов в организме вызывают его заболевание. Некоторые микроорганизмы в зависимости от условий могут существовать либо как паразиты, либо как сапрофиты. Их называют условными или факультативными паразитами. К их числу относятся возбудители многих кишечных инфекций. Другие могут развиваться только в живых клетках — это строгие, или облигатные, паразиты. Способность к биосинтезу у них самая низкая.
Для синтеза клеточных белков микроорганизмам необходим азот. По отношению к источникам азотного питания среди микроорганизмов можно выделить автоаминотрофов и гетероаминотрофов. Первые способны использовать азот неорганический (аммонийный, нитратный, молекулярный) или простейшие формы органического (мочевина) и из этих соединений строить разнообразные белки своего тела. При этом все формы азота сначала переводятся в аммонийную форму. Эта наиболее восстановленная форма азота легко трансформируется в аминогруппу. Гетероаминотрофы нуждаются в органических формах азота - белках и аминокислотах. Некоторым из них требуется полный набор аминокислот, другие создают необходимые белковые соединения из одной - двух аминокислот путем их преобразования.
Многие гетеротрофные по отношению к углероду микроорганизмы являются автоаминотрофами. К ним относятся я бактерии, участвующие в очистке сточных вод.
Потребность в кислороде и водороде для конструктивного обмена микроорганизмы удовлетворяют за счет воды и органических питательных веществ. Источниками зольных элементов (P, S, K, Mg, Fe) служат соответствующие минеральные соли. Потребность в этих элементах невелика, но присутствие в среде обязательно. Помимо того, для нормальной жизнедеятельности микробов необходимы микроэлементы – Zn, Co, Cu, Ni и др. Часть их входит в состав естественного питания микробов, часть усваивается ими из минеральных солей.
Способы получения пищи, т. е. способы питания микроорганизмов, отличаются большим разнообразием. Различают три основных способа питания: голофитное, сапрозойное, голозойное.
Голофитное питание (от греч. «голо» - целиком, «фит» - растение) совершается по типу фотосинтеза растений. Такое питание присуще только автотрофам. Среди микроорганизмов этот способ свойствен водорослям, окрашенным формам жгутиковых и некоторым бактериям.
Гетеротрофные микроорганизмы питаются либо твердыми пищевыми частицами, либо поглощают растворенные органические вещества.
Голозойное питания предопределяет развитие у микроорганизмов специальных органоидов для переваривания пищи, а у некоторых - и для ее захвата. Например, неокрашенные жгутиковые и ресничные инфузории имеют ротовое отверстие, к которому пища подгоняется соответственно жгутиками или ресничками. Наиболее высокоорганизованные инфузории образуют околоротовыми ресничками ток воды в виде воронки, направленной узким концом в рот. Пищевые частицы осаждаются на дне воронки и заглатываются инфузорией. Такие инфузории называют седиментаторами. Амебы питаются путем фагоцитоза.
Микроорганизмы с голозойным способом питания для конструктивного метаболизма используют главным образом цитоплазму других организмов - бактерий, водорослей и т. д. и имеют специальные органоиды для пищеварения. Пищеварительный процесс у простейших осуществляется в пищеварительных вакуолях.
Переваривание заключается в гидролитическом расщеплении сложных органических веществ до более простых соединений. При этом углеводы гидролизуются до простых сахаров, белки - до аминокислот, а при гидролизе липидов образуются глицерин и высшие жирные кислоты. Продукты пищеварения всасываются в цитоплазму и подвергаются дальнейшему преобразованию.
Бактерии, микроскопические грибы, дрожжи не имеют специальных органоидов для захвата пищи, и она поступает в клетку через всю поверхность. Такой способ питания называется сапрозойным.
Чтобы проникнуть в клетку, питательные вещества должны находится в растворенном состоянии и иметь соответствующий размер молекул. Для многих высокомолекулярных соединений цитоплазматическая мембрана непроницаема, а некоторые из них не могут проникнуть даже через клеточную оболочку. Однако это не означает, что высокомолекулярные соединения не используются микроорганизмами как питательные вещества. Микроорганизмы синтезируют внеклеточные пищеварительные ферменты, гидролизующие сложные соединения. Таким образом, процесс пищеварения, протекающий у простейших в вакуолях, у бактерий осуществляется вне клетки (Приложения рис. 4).
Размер молекул - не единственный фактор, обусловливающий проникновение питательных веществ в клетку.
Цитоплазматическая мембрана способна пропускать одни соединения и задерживать другие.
Известно несколько механизмов переноса веществ через мембрану клетки: простая диффузия, облегченная диффузия и Активный перенос (Приложения рис. 5).
Простая диффузия - это проникновение молекул вещества в клетку без помощи каких-либо переносчиков.
В насыщении клетки питательными веществами простая диффузия большого значения не имеет. Однако именно таким путем в клетку поступают молекулы воды. Немаловажную роль в этом процессе играет осмос - диффузия молекул растворителя через полупроницаемую перепонку в направлении более концентрированного раствора.
Роль полупроницаемой перепонки в клетке выполняет цитоплазматическая мембрана. В клеточном соке растворено огромное количество молекул разнообразных веществ, поэтому клетки микроорганизмов обладают довольно высоким осмотическим давлением. Величина его у многих микробов достигает 0,5—0,8 МПа. В окружающей среде осмотическое давление обычно ниже. Это вызывает приток воды внутрь клетки и создает в ней определенное напряжение называемое тургором.
При облегченной диффузии растворенные вещества поступают в клетку с участием специальных ферментов-переносчиков, носящих название пермеаз. Они как бы захватывают молекулы растворенных веществ и переносят их к внутренней поверхности мембраны.
Простая и облегченная диффузия представляет собой варианты пассивного транспорта веществ. Движущей силой переноса веществ в клетку в этом случае служит градиент концентраций по обе стороны мембраны. Однако большинство веществ поступает в клетку против градиента концентрации. В этом случае на такой перенос затрачивается энергия и перенос называется активным. Активный перенос протекает с участием специфических белков, сопряжен с энергетическим обменом клетки и позволяет накапливать в клетке пительные вещества в концентрации во много раз больше, чем концентрация их во внешней среде. Активный перенос - основной механизм поступления питательных веществ в клетки с сапрозойным питанием.
3. Потребность прокариот в питательных веществах
Мономеры, необходимые для построения основных клеточных компонентов, могут быть синтезированы клеткой или поступать в готовом виде из среды. Чем больше готовых соединений должен получать организм извне, тем ниже уровень его биосинтетических способностей, так как химическая организация всех свободноживущих форм одинакова.
3.1 Источники углерода
В конструктивном метаболизме основная роль принадлежит углероду, поскольку все соединения, из которых построены живые организмы, — это соединения углерода. Их известно около миллиона. Прокариоты способны воздействовать на любое известное углеродное соединение, т. е. использовать его в своем метаболизме. В зависимости от источника углерода для конструктивного метаболизма все прокариоты делятся на две группы: автотрофы, к которым принадлежат организмы, способные синтезировать все компоненты клетки из углекислоты, и гетеротрофы, источником углерода для конструктивного метаболизма которых служат органические соединения. Понятия "авто-" и "гетеротрофия" характеризуют, таким образом, тип конструктивного метаболизма. Если автотрофия — довольно четкое и узкое понятие, то гетеротрофия — понятие весьма широкое и объединяет организмы, резко различающиеся своими потребностями в питательных веществах.
Наибольшая степень гетеротрофности присуща прокариотам, относящимся к облигатным внутриклеточным паразитам, т. е. организмам, которые могут жить только внутри других живых клеток. Паразитический образ жизни привел к редукции некоторых метаболических путей у этих прокариот, что и обусловило полную их зависимость от метаболизма клетки хозяина.
Другие паразитические прокариотные организмы удается выращивать на искусственных средах, но состав таких сред необычайно сложен. Они содержат, как правило, белки или продукты их неглубокого гидролиза (пептиды), полный набор витаминов, фрагменты нуклеиновых кислот и т. д. Для приготовления питательных сред такого состава используют мясные гидролизаты, цельную кровь или ее сыворотку. Формы, способные расти при создании подходящих условий вне клетки хозяина, называют факультативными паразитами.
Следующую крупную группу прокариот составляют так называемые сапрофиты — гетеротрофные организмы, которые непосредственно от других организмов не зависят, но нуждаются в готовых органических соединениях. Они используют продукты жизнедеятельности других организмов или разлагающиеся растительные и животные ткани. К сапрофитам относится большая часть бактерий. Степень требовательности к субстрату у сапрофитов весьма различна. В эту группу входят организмы, которые могут расти только на достаточно сложных субстратах (молоко, трупы животных, гниющие растительные остатки), т. е. им нужны в качестве обязательных элементов питания углеводы, органические формы азота в виде набора аминокислот, пептидов, белков, все или часть витаминов, нуклеотиды или готовые компоненты, необходимые для синтеза последних (азотистые основания, пятиуглеродные сахара). Чтобы удовлетворить потребность этих гетеротрофов в элементах питания, их обычно культивируют на средах, содержащих мясные гидролизаты, автолизаты дрожжей, растительные экстракты, молочную сыворотку.
Есть прокариоты, требующие для роста весьма ограниченное число готовых органических соединений в основном из числа витаминов и аминокислот, которые они не в состоянии синтезировать сами, и наконец, гетеротрофы, нуждающиеся только в одном органическом источнике углерода. Им может быть какой-либо сахар, спирт, кислота или другое углеродсодержащее соединение. Описаны бактерии из рода Pseudomonas, способные использовать в качестве единственного источника углерода и энергии любое из 200 различных органических соединений, и бактерии, для которых источником углерода и энергии может служить узкий круг довольно экзотических органических веществ. Например, Bacillus fastidiosus может использовать только мочевую кислоту и продукты ее деградации, а некоторые представители рода Clostridium растут только в среде, содержащей пурины. Использовать другие органические субстраты для роста они не могут. Биосинтетические способности этих организмов развиты в такой степени, что они сами могут синтезировать все необходимые им углеродные соединения.
Особую группу гетеротрофных прокариот, обитающих в водоемах, составляют олиготрофные бактерии, способные расти при низких концентрациях в среде органических веществ. Организмы, предпочитающие высокие концентрации питательных веществ, относят к копиотрофам. Если у типичных копиотрофов оптимальные условия для роста создаются при содержании в среде питательных веществ в количестве примерно 10 г/л, то для олиготрофных организмов — в пределах 1—15мг углерода/л. В средах с более высоким содержанием органических веществ такие бактерии, как правило, расти не могут и погибают.
3.2 Азот
Азот является одним из четырех основных элементов, участвующих в построении клетки. В расчете на сухие вещества его содержится приблизительно 10%. Природный азот бывает в окисленной, восстановленной и молекулярной формах. Подавляющее большинство прокариот усваивают азот в восстановленной форме. Это соли аммония, мочевины, органические соединения (аминокислоты или пептиды). Окисленные формы азота, главным образом нитраты, также могут потребляться многими прокариотами. Так как азот в конструктивном клеточном метаболизме используется в форме аммиака, нитраты перед включением в органические соединения должны быть восстановлены.
Восстановление нитратов до аммиака осуществляется посредством последовательного действия двух ферментов — нитрат- и нитритредуктазы.
Давно была обнаружена способность отдельных представителей прокариотного мира использовать молекулярный азот атмосферы. В последнее время показано, что этим свойством обладают многие прокариоты, принадлежащие к разным группам: эу- и архебактерии, аэробы и анаэробы, фототрофы и хемотрофы, свободноживущие и симбиотические формы. Фиксация молекулярного азота также приводит к восстановлению его до аммиака.
3.3 Потребности в источниках серы и фосфора
Сера входит в состав аминокислот (цистеин, метионин), витаминов и кофакторов (биотин, липоевая кислота, кофермент А и др.), а фосфор — необходимый компонент нуклеиновых кислот, фосфолипидов, коферментов. В природе сера находится в форме неорганических солей, главным образом сульфатов, в виде молекулярной серы или входит в состав органических соединений. Большинство прокариот для биосинтетических целей потребляют серу в форме сульфата, который при этом восстанавливается до уровня сульфида. Однако некоторые группы прокариот не способны к восстановлению сульфата и нуждаются в восстановленных соединениях серы. Основной формой фосфора в природе являются фосфаты, которые и удовлетворяют потребности прокариот в этом элементе.
3.4 Необходимость ионов металлов
Всем прокариотным организмам необходимы металлы, которые могут использоваться в форме катионов неорганических солей. Некоторые из них (магний, кальций, калий, железо) нужны в достаточно высоких концентрациях, потребность в других (цинк, марганец, натрий, молибден, медь, ванадий, никель, кобальт) невелика. Роль перечисленных выше металлов определяется тем, что они входят в состав основных клеточных метаболитов и, таким образом, участвуют в осуществлении жизненно важных функций организма.
3.5 Потребность в факторах роста
Некоторые прокариоты обнаруживают потребность в одном каком-либо органическом соединении из группы витаминов, аминокислот или азотистых оснований, которое они по каким-то причинам не могут синтезировать из используемого источника углерода. Такие органические соединения, необходимые в очень небольших количествах, получили название факторов роста. Организмы, которым в дополнение к основному источнику углерода необходим один или больше факторов роста, называют ауксотрофами, в отличие от прототрофов, синтезирующих все необходимые органические соединения из основного источника углерода.
4. Типы метаболизма микроорганизмов
Для полной характеристики микроорганизмов используют понятие тип метаболизма. Различия в типах метаболизма определенных групп микроорганизмов обусловлены особенностями конструктивного и спецификой энергетического обменов. В зависимости от используемого источника энергии для получения АТФ микроорганизмы делят на фототрофов (используют энергию света) и хемотрофов (используют энергию химических реакций).
Процесс образования АТФ называется фосфорилированием; он осуществляется в митохондриях (у эукариот) и ферментных системах, локализованных на цитоплазмаческой мембране (у прокариот). Механизм образования, АТФ у разных групп микроорганизмов неодинаков. Различают субстратное, окислительное и фотофосфорилирование. Любой тип фосфорилирования обязательно сопряжен с переносом электронов в ходе окислительно-восстановительных реакций энергитического обмена. При этом одни микроорганизмы в качестве доноров электронов (водорода) используют неорганические, другие - органические соединения. Соответственно первые называются литотрофами, вторые - органотрофами.
Таким образом, принимая во внимание тип питания (авто- или гетеротрофное), природу донора электронов источник энергии (свет или химическая реакция), возможные сочетания вариантов конструктивного и энергетического обменов можно представить в виде следующей схемы.
Каждый из представленных вариантов характеризует определенный тип метаболизма. В табл. 1 приведены представители микроорганизмов каждого типа метаболизма
Большинство микроорганизмов, обитающих в природных источных водах и играющих важную роль в формировавании качества воды и ее очистке, относятся к восьмому и первому типам метаболизма. В связи с этим при дальнейшем изложении материала именно им уделено основное внимание.
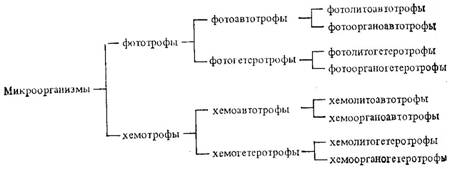 |
Схема 1. Варианты конструктивного и энергетических обменов.
5. Энергетический метаболизм фототрофов
Все указанные в табл. 1 фотосинтезирующие микроорганизмы приспособлены к использованию света видимого (длинна волны 400—700 нм) и ближней инфракрасной части спектра (700—1100 нм). Эта способность существовать за счет энергии света обусловлена присутствием в клетках органоидов со специфическими светочувствительными пигментами. Каждому виду микроорганизмов свойствен характерный и постоянный набор пигментов.
Таблица 1
| Тип метаболизма | Представители |
| 1) Фотолитоавтотрофия | Водоросли, цианобактерии, большинство пурпурных бактерий и зеленых серобактерий. |
| 2) Фотолитогетеротрофия | Частично цианобактерии пурпурные и зеленые серобактерии |
| 3) Фотоорганоавтотрофия | Некоторые пурпурные бактерии |
| 4) Фотоорганогетеротрофия | Большинство несерных пурпурных бактерий |
| 5) Хемолитоавтотрофия | Нитрифицирующие, тионовые, некторые железобатерии. |
| 6) Хемолитогетеротрофия | Бесцветные серобактерии |
| 7) Хемоорганоавтотрофия | Некоторые бактерии окисляющие муравьиную кислоту |
| 8) Хемоорганогетеротрофия | Простейшие, грибы, большинство бактерий. |
Для некоторых представителей группы цианобактерий наряду с фотолитоавтотрофией показана способность к фотолито- или хемоорганогетеротрофии. Ряд хемолитоавтотрофных видов Thiobacillus способы существовать за счет использования в качестве источников энергии и углерода органических соединений, т. е. хемоорганогетеротрофно.
Некоторые прокариоты могут существовать только на базе одного какого-нибудь способа питания. Например, одноклеточная цианобактерия Synechococcus elongatus может использовать в качестве источника энергии только свет, а как основной источник углерода в конструктивном метаболизме — углекислоту. Характеризуя способ существования (образ жизни, тип метаболизма) этого организма, мы говорим, что он облигатный фотолитоавтотроф. Многие бактерии, относящиеся к роду Thiobacillus, — облигатные хемолитоавтотрофы, т. е. источником энергии для них служат процессы окисления различных соединений серы, а источником углерода для построения веществ тела, — углерод углекислоты. Подавляющее большинство бактерий — облигатные хемоорганогетеротрофы, использующие в качестве источника углерода и энергии органические соединения.
Световая энергия улавливается системой поглощающих пигментов и передается в реакционный центр, который возбуждает молекулы хлорофилла. В темноте молекула хлорофилла находится в стабильном невозбужденном состоянии, когда свет падает на эту молекулу, она возбуждается и один из электронов на более высокий энергитический уровень. Молекулы хлорофилла тесно связаны с системой транспорта электронов. Каждый квант поглощенного света обеспечивает отрыв от молекулы хлорофилла одного электрона, который, проходя по цепи переноса электронов, отдает свою энергию системе АДФ-АТФ, в результате чего энергия света трансформируется в энергию макроэргической связи молекулы АТФ. Такой способ образования АТФ называется фотосинтетическим фосфорилированием.
Однако для осуществления биосинтетических процессов продуктивного обмена микроорганизмам кроме энергии необходим восстановитель - донор водорода (электронов). Для водорослей и цианобактерий таким экзогенным донором водорода служит вода. Восстановление диоксида углерода в процессе фотосинтеза и превращение его в структурные компоненты клетки у этих видов микроорганизмов протекает аналогично фотосинтезу высших растений:
СО2+Н2О→(СН2О)+О2
Формула СН2О символизирует образование органического соединения, в котором уровень окисленности углерода примерно соответствует окисленности углерода в органических веществах клетки.
У фотосинтезирующих бактерий донорами водорода реакций синтеза могут быть как неорганические, так органические вещества. Большинство пурпурных и зеленых серобактерий, относящихся к группе фотолитоавтоавтотрофов восстанавливает СО2, используя Н2S как донор водорода:
СО2+2Н2S→(CH2O)+H2O+2S
Такой тип фотосинтеза получил название фоторедукцищ Основное отличие бактериальной фоторедукции от фотосинтеза и зеленых растений и водорослей заключается в том, что донором водорода служит не вода, а другие соединения и фоторедукция не сопровождается выделением кислорода.
В отличие от неорганических восстановителей, которые выполняют роль только доноров водорода, экзогенные органические восстановители могут одновременно служить и источниками углерода (фотоорганогетеротрофия).
Способность использовать органические соединения той или иной степени присуща всем фотосинтезирующим бактериям. Для фотолитогетеротрофов они служат только источниками углеродного питания, для фотоорганоавтотрофов - только донорами водорода. Например, несерные пурпурные бактерии рода Rhodopseudomonas sp. могут осуществлять фотосинтез, используя в качестве донора водорода изопропанол, восстанавливая при этом диоксид углерода и продуцируя ацетон:
энергия АТФ
СО2 +2СН3СНОНСН3→(СН2О)+ 2СН3СОСН3 +Н2О
6. Энергетический метаболизм хемотрофов, использующих процессы брожения
Из трех путей образования АТФ субстратное фосфорилиронание наиболее простой. Такой тип энергетического метаболизма характерен для многих бактерий и дрожжей, осуществляющих различные виды брожения.
Брожение идет в анаэробных условиях и может быть определено как процесс биологического окисления сложных органических субстратов для получения энергии, при котором конечный акцептор водорода (также органическое вещество) образуется в ходе распада исходного субстрата. При этом одни органические вещества служат донорами водорода и окисляются, другие - акцепторами водорода и в результате восстанавливаются. Перенос водорода от доноров к акцепторам осуществляется с помощью окислительно - восстановительных ферментов.
Кроме углеводов многие бактерии способны сбраживать самые разнообразные соединения: органические кислоты, аминокислоты, пурины и т. д. Условие, определяющее способность вещества к сбраживанию, - наличие в его структуре не полностью окисленных (восстановленных) атомов углерода. Только в этом случае возможна внутри - и межмолекулярная перестройка субстрата за счет сопряжения окислительных и восстановительных реакций без участия кислорода.
В результате процессов брожения в среде накапливаются вещества, в которых степень окисления углерода может быть как выше, так и ниже, чем в исходном субстрате. Однако строгое равновесие окислительных и восстановительных процессов при брожении приводит к тому, что средняя степень окисления углерода остается такой же, как и у субстрата.
Существует несколько типов брожений, названия которым даются по конечному продукту: спиртовое (осуществляют дрожжи и некоторые виды бактерий), пропионовокислое (пропионовые бактерии), метановое (метанобразующие бактерии), маслянокислое (маслянокислые бактерии) и т.д.
Многие микроорганизмы, осуществляющие процессы брожения,- облигатные анаэробы, не способные развиваться в присутствии кислорода и даже более слабых окислителей. Другие - факультативные анаэробы - могут расти как в кислородной среде, так и в бескислородной. Это отличительное свойство факультативных анаэробов объясняется тем, что они могут изменять способ образования АТФ переключаться с окислительного фосфорилирования при наличии в среде кислород на субстратное его отсутствии. Характерная особенность процессов биологического окисления - их многостадийность. обеспечивающая постепенное выделение свободной энергии, заключенной в сложных органических субстратах.
Многостадийность энергетического метаболизма принципиально необходима для жизнедеятельности любого организма. Если бы в клетке окисление сложных веществ протекало в одну стадию, то одновременное освобождение нескольких сотен килоджоулей привело бы к выделению большого количества тепла, резкому повышению температуры и к гибели клетки, поскольку эффективность использования энергии ограничена возможностями системы АДФ—АТФ.
Простейший пример анаэробного окисления глюкозы - молочнокислое брожение. Оно вызывается молочнокислыми бактериями, факультативными анаэробами, не образующими спор. Превращение ПВК при молочнокислом брожении протекает следующим образом:
СН3СОСООН + НАД*Н2, — СН3СНОНСООН + НАД
Значительно сложнее механизм пропионовокислого брожения, служащего источником энергии группе пропионовых бактерий, факультативных анаэробов, неподвижных не спорообразующих бактерий рода Propionibacterium. Эти бактерии синтезируют конечный акцептор, присоединяя к молекуле ПВК СО2. Процесс известен под названием гететеротрофной ассимиляции СО2. В результате образуется щавелевоуксусная кислота - акцептор водорода для НАД*Н2. Дальнейшие ферментативные реакции приводят к образованию пропионовой кислоты.
Маслянокислое брожение осуществляют бактерии род Clostridium. Таким образом, энергетический выход процесса брожения невелик, поскольку органические вещества не окисляются полностью и часть энергии исходного субстрата сохраняется в достаточно сложных продуктах брожения. В большинстве случаев при сбраживании глюкозы клетка запасает две молекулы АТФ на 1 моль глюкозы.
Для получения энергии, необходимой для синтеза клечного вещества и других жизненных функций, микроорганизмам, осуществляющим процессы брожения, приходится перерабатывать большое количество органических веществ.
Именно в силу этих причин на очистных станциях систем водоотведения анаэробные процессы брожения используют для обработки концентрированных субстратов – осадков сточных вод.
Энергетический метаболизм хемоорганотрофов, использующих процесс дыхания.
Большинство гетеротрофных организмов получают энергию в процессе дыхания - биологического окисления сложных органических субстратов, являющихся донорами водорода. Водород от окисляемого вещества поступает в дыхательную цепь ферментов. Дыхание называют аэробным, если роль конечного акцептора водорода выполняет свободный кислород. Микроорганизмы, способные существовать только в присутствии кислорода, называются облигатными аэробами.
В качестве источников энергии - доноров водорода -хемоорганогетеротрофы в процессе дыхания могут использовать разнообразные окисляемые органические соединения: углеводы, жиры, белки, спирты, органические кислоты и т. д. Суммарно процесс дыхания при окислении углеводов выражается следующим уравнением:
С6Н12О6 + 6О→ 6СО2 + 6Н2О + 2820 кДж
Начальная стадия превращения углеводов вплоть до образования ПВК полностью идентична ферментативным реакциям окисления углеводов в процессе брожения.
В клетках аэробов ПВК может быть окислена полностью в результате ряда последовательных реакций. Совокупность этих превращений составляет цикл, именуемый циклом Кребса или циклом ди- и трикарбоновых кислот (ЦТК).
Водород, отнятый дегидрогеназами в цикле, передается в дыхательную цепь ферментов, которая у аэробов кроме НАД включает ФАД, систему цитохромов и конечный акцептор водорода - кислород. Передача водорода по этой цепи сопровождается образованием АТФ.
Первый этап фосфорилирования связан с пере