


Отдел красные водоросли
ОТДЕЛ КРАСНЫЕ ВОДОРОСЛИ
В хроматофорах красных водорослей, помимо хлорофиллов и каротиноидов (р-каротин, зеаксантин, антераксантин, криптоксантин, лютеин, неоксантин), содержатся еще водорастворимые пигменты — фикобилины: фикоэритрины красного цвета, фикоцианины и аллофикоциан синего цвета. От соотношения пигментов зависит окраска таллома, варьирующая от малиново-красной до голубовато-стальной. Хлоропласты одеты оболочкой из двух мембран и содержат одиночные тилакоиды, на поверхности которых локализованы фикобилисомы. Генофор рассеянный. Периферические тилакоиды у более примитивных форм со звездчатыми или сильно лопастными хроматофорами с пиреноидами отсутствуют, а у более высокоорганизованных красных водорослей с дисковидными хлоропластами без пиреноидов имеются. Запасной продукт — полисахарид «багрянковый крахмал», от йода приобретающий буро-красный цвет. Зерна багрянкового крахмала откладываются в цитоплазме всегда вне связи с пиреноидами и хроматофорами. Для жизненного цикла характерно полное отсутствие жгутиковых стадий и особая форма моногамного полового процесса. Как и бурые, красные водоросли — почти исключительно морские.
Строение таллома красных водорослей довольно простое: известны даже одноклеточные коккоидные формы, у многих талломы гетеротрихальные и в виде разветвленных нитей, прикрепленных к субстрату с помощью ризоидов. Однако подавляющее большинство имеет псевдопаренхиматозные талломы, возникающие за счет переплетения и срастания боковых ветвей либо одной оси, неограниченно нарастающей с помощью верхушечной клетки, либо многих таких осей; в первом случае говорят об одноосевом строении, во втором — о многоосевом или мультиосевом. Пластинчатые талломы истинно паренхиматозного строения, получающиеся в результате как поперечных, так и продольных делений клеток, встречаются среди красных водорослей редко.
Клетка красных водорослей одета оболочкой, пектиновые и темицеллюлозные компоненты которой сильно набухают и часто сливаются в общую слизь мягкой или хрящеватой консистенции, заключающей протопласты. Нередко в оболочках откладывается известь. При делении клеток у большинства красных водорослей в результате неполного смыкания центрипетально растущей поперечной стенки образуются первичные поры. Поровый канал сначала открыт, но затем замыкается пробкой. По этим порам можно восстановить генетическую связь между клетками в многоклеточном талломе. Клетки одно- и многоядерные, хроматофоры, как правило, париетальные, многочисленные, в виде зерен или пластинок. У бангиофициевых встречаются звездчатые хроматофоры с центральными пиреноидами.
Размножение. Вегетативное размножение — путем фрагментации таллома; особенно часто наблюдается в местообитаниях, где факторы внешней среды неблагоприятны для размножения спорами.
Бесполое размножение осуществляется посредством неподвижных клеток, развивающихся из содержимого спорангия в числе одной или четырех.
Тетраспоры формируются на диплоидных бесполых растениях—тетраспорофитах. В тетраспорангиях перед образованием тетраспор происходит мейоз. Расположение тетраспор в тетраспорангии может быть крестообразным, рядовым или по углам тетраэдра.
При крестообразном расположении содержимое тетраспорангия сначала делится в горизонтальном направлении, а затем каждая половина, в свою очередь, делится вертикально. При рядовом или зональном расположении тетраспор содержимое вытянутого тетраспорангия делится поперечными перегородками на четыре клетки, расположенные друг над другом. Если тетраспоры расположены по углам тетраэдра, содержимое тетраспорангия сразу распадается па 4 тетраспоры. Гаплоидные тетраспоры прорастают в гаплоидные растения — гаметофиты того же или иного, чем тетраспорофиты, облика. Они размножаются половым путем: на них развиваются половые органы — гаметангии.
Половой процесс оогамный. Женский орган – карпогоп — у большинства красных водорослей состоит из расширенной базальной части — брюшка, заключающего яйцеклетку, и отростка — трихогины. Трихогина или вообще не содержит ядра, или оно дегенерирует к моменту оплодотворения. Карпогон обычно развивается на особой короткой, состоящей из 3—4, реже из 7—9 клеток карпогонной ветви, которая в свою очередь сидит на поддерживающей клетке. Антеридии — обычно мелкие бесцветные клетки, содержимое которых освобождается в виде мелких голых, лишенных жгутиков мужских оплодотворяющих элементов— спермациев. Выпавшие из антеридиев спермации пассивно переносятся токами воды и прилипают к трихогине. В месте контакта спермация и трихогины их стенки растворяются, и ядро спермация по трихогине перемещается в брюшную часть карпогона, где сливается с женским ядром. Эти данные классических исследований конца прошлого века, сохранившие значение до наших дней, можно дополнить некоторыми наблюдениями последних лет. Так, в 1984 г, получены новые данные о динамике полового процесса у некоторых красных водорослей. В условиях эксперимента у Caliithamnion cordatum удалось показать, что зрелые карпогоны восприимчивы к спермациям в течение 3 ч. Сам процесс слияния гамет, включающий известные из классических работ фазы, длится 5—10 ч. После оплодотворения базальная часть карпогона отделяется перегородкой от трихогииы, которая отмирает, и претерпевает дальнейшее развитие, приводящее у большинства красных водорослей к образованию карпоспор. Детали этого развития имеют важное систематическое значение. У одних красных водорослей содержимое зиготы непосредственно делится с образованием неподвижных голых спор карпоспор, у других из оплодотворенного карпогона вырастают многоклеточные ветвящиеся нити — гонимоблаапы, клетки которых превращаются в карпоаюрангии, производящие по одной карпоспоре. Наконец, у большинства красных водорослей гонимобласты развиваются не непосредственно из брюшка оплодотворенного карпогона, а из особых вспомогательных — ауксилярных клеток. Последние могут быть удалены от карпогона или же располагаются на талломе в непосредственной от него близости. Если ауксилярные клетки удалены от карпогона, из его брюшка после оплодотворения вырастают гифообразные, без поперечных клеточных перегородок соединительные или ообластемные нити. Их возникновению предшествуют митотические деления копуляционного ядра и таким образом они содержат диплоидные ядра. Ообластемные нити подрастают к ауксилярным клеткам, в точке контакта оболочки растворяются, и кончик ообластемной нити и аукенлярная клетка сливаются. Диплоидное ядро ообластемной нити делится, одно из дочерних ядер передается ауксилярной клетке, а другое остается в ообластемной нити, которая может расти к следующей ауксилярной клетке, и т. д. Таким образом, все большее число ауксилярных клеток обеспечивается диплоидным ядром — потомком диплоидного копуляционного ядра одного-единственного оплодотворенного карпогона. Слияние кончика ообластемной нити с ауксилярной клеткой не сопровождается слиянием их ядер — диплоидного ообластемной нити и гаплоидного ауксилярной клетки. Диплоидизированная ауксилярная клетка, отделяясь перегородкой от ообластемной нити, делится на базальную клетку и центральную. Первая получает первоначальное гаплоидное ядро ауксилярной клетки, вторая—диплоидное ядро и становится центром развития гонимобластов, клетки которых содержат диплоидные ядра и продуцируют диплоидные карпоегюры. Гонимобласты, развивающие карпоспоры, рассматриваются как особая генерация — карпоспорафит.
У вышестоящих порядков красных водорослей ауксилярные клетки формируются рядом с карпогоном до оплодотворения карпогона или только после того, как оно произошло. Совокупность ауксилярной клетки с карпогоном называется прокарпием. Здесь нет необходимости в образовании длинных ообластемных нитей, связывающих карпогоп и ауксилярные клетки, ауксилярная клетка просто сливается с брюшком оплодотворенного карпогона, после чего из нее развиваются гонимобласты с карпоспорами. Карпоспорангии часто располагаются тесными группами — цисто-карпиями, которые у многих представителей одеты псевдопаренхиматозной оболочкой, развивающейся из соседних с карпогоном клеток.
Жизненные циклы красных водорослей отличаются большим разнообразием, что в последнее время было продемонстрировано на основе следующих данных, полученных разными методами. Наблюдения в природе дают представление о встречаемости морфологических фаз и репродуктивных органов, на них развивающихся. В условиях лабораторной культуры можно проследить прорастание репродуктивных клеток и развитие из них талломов до состояния, когда они дают репродуктивные органы, т. е. в культуре можно строго установить последовательность морфологических фаз. Особенно плодотворным оказался метод лабораторных культур для установления связей между гаметангиальной и тетраспорангиальной фазами, различающимися морфологически и долгое время рассматриваемыми как самостоятельные таксоны. Цитологический метод позволяет установить число хромосом каждой морфологической фазы, место в жизненном цикле сингамии, мейоза и других цитологических деталей. Ниже приводятся жизненные циклы, обоснованные совокупностью вышеперечисленных данных,
У красных водорослей наиболее распространен жизненный цикл, состоящий в последовательности гаметангиальной, карпоспорангиальной и тетраспорангиальной фаз, причем первая и последняя морфологически сходны, а карпогониальная фаза развивается на гаметангиальной. Такая изоморфная смена гаметофитов и тетраспорофитов впервые была описана для Polysiphonia violaceae. Поэтому такой жизненный цикл, присущий большинству красных водорослей, обычно называют Polysiphonia-тип. Его можно встретить помимо порядка церамиальных, куда относится род полисифония, и у представителей порядков криптонемиальные, гигартинальные, родимениальные и гелидиальные.
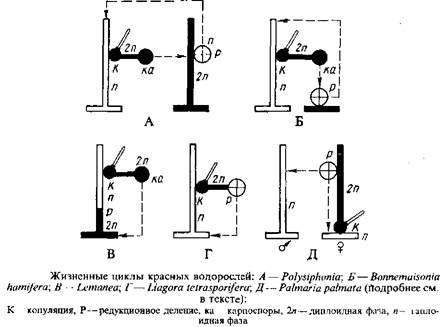
Предположение о жизненном цикле Polysiphonia-типа у представителей вышестоящих порядков класса флоридеофициевых в последние годы подтверждено строгими культуральными экспериментами.
Цикл развития Bonnemaisonia hamifera-тина, который можно назвать гетероморфной сменой генераций, характеризуется последовательностью гаметанги-альной, карпоспорапгиальной и тетраспорангиальной фаз, причем все три фазы морфологически неодинаковы. Карпогониальная фаза развивается на гаметангиальной. В последние годы такой жизненный цикл был описан не только для многих представителей порядка немалиальных, но и для криптонемиальных и гигартинальньгх.
У Liagora tctrasporifera в жизненном цикле чередуются гаметангиальная и тетраспорангиальная фазы, морфологически различающиеся, причем тетраспорангиальная фаза развивается на гаметангиальной; мейоз осуществляется в тетраспорангиях, карпоспорофит отсутствует.
У Palmaria paimata также отсутствует карпоспорофит и наблюдается чередование гаметангиальной и тетраспорангиальной фаз, причем вторая развивается на первой. Однако женский гаметофит здесь крохотный, а мужской — крупный, морфологически неотличимый от тетраспорофита.
Приведенные жизненные циклы характеризуются спорической редукцией, так как мейоз происходит при образовании тетраспор.
Жизненный цикл, описанный пока только для двух родов — леманеа и батрахоспермум, отличается соматической редукцией. В апикальной клетке предростка, развивающегося из диплоидных карпоспор, происходит мейоз и из нее вырастает гаплоидная гаметангиальная фаза.
Сказанным не исчерпывается все разнообразие жизненных циклов красных водорослей.
Красные водоросли делятся на два класса: бангиофициевые и флоридеофициевые
КЛАСС БАНГИОФИЦИЕВЫЕ — BANGIOPHYCEAE
Клетки часто со звездчатым хроматофором, заключающим пиреноид. Поры между клетками, как правило, отсутствуют. Кар-погон без трихогины, после оплодотворения содержимого карпогона непосредственно делится с образованием карпоспор. Бесполое размножение обычно моноспорами.
Представителями могут служить роды порфира и бангия. Род порфира характеризуется листоватыми талломами истинно паренхиматозного строения, прикрепленными своими основаниями к субстрату и достигающими 50 см в длину, редко более. Пластинка состоит из одного слоя клеток или двухслойная. Клетки содержат по одной или по две пластиды. Пластинки у разных видов неодинаковой формы с цельными или зазубренными краями, различной окраски и толщины. Проростки сначала представлены однорядными нитями. Последовательные продольные деления клеток этих нитей приводят к образованию листовидных талломов. Порфира распространена как в южных, так и в северных морях в прибрежной литоральной зоне.
Род бангия, включающий как морские, так и пресноводные виды, имеет таллом, представленный неразветвленными нитями, вначале прикрепленными к субстрату базальной клеткой; с возрастом от нижних клеток развиваются песептированные ризоиды. Нити сначала однорядные, но в более старых участках клетки претерпевают продольные радиальные деления, так что на поперечном срезе таллома видны сектора.
Зрелые талломы порфиры и бангии размножаются половым путем. У порфиры при образовании антеридиев клетки талломов делятся во взаимно перпендикулярных направлениях на ряд мелких клеток, каждая из которых образует по одному спермацию. При разбухании интерцеллюлярной слизи спермации выдавливаются и освобождаются. Карпогоны лишь незначительно отличаются от вегетативных клеток. Типичная трихогина отсутствует. После оплодотворения зигота непосредственно делится на 2—32 карпоспоры, которые освобождаются так же. как и спермации,— благодаря набуханию межклеточной слизи. Сходным образом осуществляется половое размножение бангии. Так. Е. Мань утверждает, что талломы бангии гаплоидны и представляют собой гаметофиты. Они двудомны: мужские талломы продуцируют спермации, которые копулируют с клетками женских нитей, функционирующими как каргюгоны. В результате возникает диплоидная зигота, которая делится на карпоспоры.
Цикл развития. Цикл развития порфиры и бангии интенсивно изучался в последние десятилетия как в природе, так и в искусственных культурах. Для многих видов порфиры и бангии в их жизненном цикле были установлены Conchocelis-стадии: карпоспоры, сначала голые, выделяют оболочку и прорастают униполярно в нитчатые растеньица, образующие на поверхности и внутри известковых раковинок моллюсков розовые пятна1. Эта стадия долгое время считалась самостоятельной водорослью Conchoceiis rosea. Нити конхоцелиса сложены из удлиненных клеток с париетальным хроматофором. Ультраструктура клеток типична для красных водорослей из класса флоридеофициевых: слоистая клеточная оболочка; одиночные тилакоиды в хлоропластах, окруженных двухмембранной хлоропластной оболочкой: периферические тилакоиды: пиреноид, в который проникают одиночные тилакоиды; рассеянные между тилакоидами фибриллы ДНК; в цитоплазме — гранулы багрянкового крахмала. Между соседними клетками Conchocelis-нитей имеются закупоренные пробками поры, сходные с таковыми в клетках флоридеофициевых.
В деталях Conchocelis-стадии разных видов порфиры и бангии обнаруживают постоянные различия: морфологические, ультраструктурные, касающиеся числа и расположения тилакоидов, пронизывающих пиреноиды, пор и замыкающих их пробок и др. Имеются постоянные различия в характере росла в культуре и физиологические, в частности, каждый вид по-своему реагирует на температуру и фотопериод. Зрелые Conchocelis-нити образуют специализированные веточки — конхоспорангиальные ветви, сложенные из изодиаметрических толстостенных клеток со звездчатыми осевыми хроматофорами без периферических тилакоидов. Клетки конхоспорангиальных ветвей освобождают конхоспоры, прорастающие биполярно и образующие проростки, которые вырастают в новые макроскопические листовые талломы. Для некоторых видов было показано, что карпоспоры и вегетативные клетки конхоцелис-стадии диплоидны. Ввиду дип-лоидности конхоцелис-стадия может быть сравнима с тетраспорофитом, хотя она и не образует тетраспор. Прежние утверждения, что мейоз у порфиры происходит при прорастании зиготы, не подтвердились, так же как и у так называемых гаплобионтных немалиальньгх. Для P. gardneri описано оплодотворение карпогона спермацием и высказывалось предположение, что мейоз имеет место при образовании конхоспор. Недавние цитологические исследования P. yezoensis дают возможность предположить, что мейоз осуществляется при образовании проростков из конхоспор. Однако у очень многих видов цитологические детали жизненного цикла неизвестны.
Каждая из морфологических фаз способны к саморепродукции. Так, молодые талломы P. yezoensis, P. tenera продуцируют обильные моноспоры, а у Р. ки-niedai моноспоры образуются обильно на протяжении всего периода жизни, даже во взрослых талломах, которые формируют спермации и карпоспоры. Прорастая, моноспоры дают листоватые талломы порфиры. Однако у P. pseudolinearis размножение посредством моноспор отсутствует. Наоборот, некоторые виды порфиры размножаются только бесполым путем — посредством аплано-спор, образующихся по 8—32 в клетках по краю таллома. Эти споры имеют звездчатый хлоропласт с центральным пиреноидом и прорастают биополярно, давая начало новому листоватому таллому.
На Conchocelis-нитях в качестве одиночных боковых ветвей возникают моноспорангии. Они снабжены ножками или сидячие, как у Bangia fuscopurpurea, располагаются поодиночке {P. miniata) или образуют цепочки. Униполярно прорастающие моноспоры вновь дают Conchocelis-стадию. Помимо моноспор, Conchocelis-нити могут размножаться вегетативно — фрагментацией таллома. При культивировании некоторых видов порфиры в качестве посевного материала используются культивируемые Conchocelis-нити.
Ряд авторов поставили под сомнение даже оплодотворение спермацием карпогона и образование карпоспор как прямого следствия полового процесса. Это повлекло к изменению терминологии. Например, карпоспоры и спермации были обозначены соответственно как гх- и р-споры. а-Споры Прорастают непосредственно в Conchoceiis-стадию, предложил классификацию спор, обнаруживаемых в цикле развития бангия и порфира, с учетом в первую очередь характера их прорастания и их дальнейшей судьбы. Он различает споры, прорастающие униполярно, биполярно и не прорастающие совсем. К униполярно прорастающим спорам относятся, например, споры, образующиеся путем повторного деления материнских клеток макроскопического таллома на 16 или менее частей и которые при определенных условиях освещения дают начало коихоцелис-фазе. Униполярно прорастают также моноспоры, репродуцирующие конхоцелис-стадию. К спорам, прорастающим биполярно, относятся моноспоры, возникающие на макроскопическом талломе, и необходимые для репродукции этой фазы, конхоспоры, развивающиеся на Conchocelis-нитях и дающие начало макроскопической фазе, а также апланоспоры, репродуцирующие у некоторых видов порфиры листоватые талломы. Для прорастания всех этих спор требуется определенный фотопериод. Споры, не прорастающие, образуются путем повторных делений содержимого'материнской клетки па более чем (6 продуктов деления.
Порфира — важная промысловая водоросль, она высоко ценится как продукт питания с высоким содержанием протеина и витаминов, а также применяется в медицине.
КЛАСС ФЛОРИДЕОФИЦИЕВЫЕ — FLORIDEOPHYCEAE
Клетки большей частью с париетальными хроматофорами без пиреноидов. Между клетками имеются поры. Карпогон с трихогиной. После оплодотворения развиваются гонимобласты или непосредственно из брюшка оплодотворенного карпогона, или из ауксилярных клеток после слияния их с ообластемными нитями или оплодотворенным карпогоном. Бесполое размножение большинства— с помощью тетраспор.
Делали развития карпоспорофита, а также время дифференцировки ауксилярных клеток, их положение на талломе и т. п. положены в основу деления флоридеофициевых на порядки.
Порядок немалиальные —nemaliales
Этот порядок, включающий формы одноосевого и многоосевого строения, характеризуется отсутствием ауксилярных клеток; гонимобласты развиваются непосредственно из оплодотворенного карпогона или из его дочерней клетки. Долгое время считалось, что у представителей этого порядка мейоз совершается непосредственно после оплодотворения при прорастании зиготы и называли их гаплобионтными флоридеями, противопоставляя всем остальным диплобионтным порядкам флоридей, у которых мейоз происходит при образовании тетраспор. Однако Е. Мань продемонстрировал цитологически отсутствие мейоза в карпогоне вслед за оплодотворением и диплоидность гонимобластов, карпоспор и продуктов их прорастания для ряда представителей порядка немалиальных. Он выдвинул гипотезу существования у них спорофитной генерации или ее эквивалента, где обязательно должен происходить мейоз. Исследования двух последних десятилетий подтвердили гипотезу, выдвинутую Е. Мань, и, кроме того, обнаружили заметное разнообразие жизненных циклов в пределах этого порядка. Здесь можно различить по крайней мере три типа циклов развития, хорошо обоснованных как с точки зрения последовательности морфологических фаз, так и цитологических данных о числе хромосом в каждой морфологической фазе и месте сингамии и мейоза в жизненном цикле. Первый тип цикла развития — тип Lemanea. В этом типе две морфологические фазы совмещены в одной структуре, где нижняя базальная часть диплоидна, а верхняя часть гаплоидна и представляет собой гаметангиальную фазу, на которой развивается карпо-спорангиальная фаза. Мейоз происходит соматически во время развития этой структуры.
Как уже отмечалось, этот тип развития установлен пока только для родов леманеа и батрахоспермум.
Род леманеа встречается в быстро текущих речках с холодной водой. Таллом, имеющий вид неветвящейся щетинки 10—15 см длиной и 1 мм толщиной, темно-фиолетового или оливково-бурого цвета с узловатыми вздутиями; прикрепляется к субстрату при помощи подошвы из стелющихся нитей. По оси таллома проходит одна нить из вытянутых бесцветных клеток. От верхней части каждой клетки центральной оси отходит мутовка обычно из четырех расходящихся по радиусам ветвей. Базальные клетки каждой такой ветви крупные, вытянутые. От их дистального конца отходят ветви второго порядка, которые, в свою очередь, многократно ветвятся. Конечные разветвления срастаются в многослойную кору. Ее наружные клетки мелкие и заполнены хроматофорами, внутренние — крупные и почти бесцветные. От проксимальных концов основных клеток радиальных ветвей отходят коровые нити, окутывающие центральную нить. Антеридии образуются группами из поверхностных клеток коры. Карпогонные ветви, несущие карпогоны, возникают от внутренних клеток коры. Брюшко карпогона находится около внутренней поверхности коры, а трихогина прорывает кору и высовывается наружу. После оплодотворения из брюшной части карпогона развиваются пучки гонимобластов, врастающие в полость таллома. Карпоспорангии формируются целыми четками, карпоспоры скапливаются внутри полости таллома и освобождаются после разрушения коры таллома. Согласно исследованиям Е. Мань, при развитии из зиготы гонимобластов мейоз не осуществляется и, таким образом, карпоспорофит и карпоспоры диплоидны. Карпоспоры прорастают с образованием диплоидного нитчатого, состоящего приблизительно из 20 клеток предростка, ядро в верхушечной клетке которого редукционно делится. Из апикальной клетки после мейоза развивается гаплоидное растение — гаметофит леманеи; на нем образуются половые органы. Таким образом, базальная часть таллома леманеи диплоидна.
У рода батрахоспермум цикл развития такой же, как у леманеи. Его представители встречаются в реках с чистой прозрачной водой и озерах. Таллом имеет вид сильно разветвленного, слизистого на ощупь кустика оливково-зеленого или стального цвета. Ось, неограниченно нарастающая за счет деятельности верхушечной клетки, состоит из одного ряда вытянутых бесцветных клеток. От каждой из них непосредственно под поперечой перегородкой возникает мутовка ветвей ограниченного роста, которые сложены из мелких, богатых хроматофорами клеток. Конечные клетки ветвей могут вытягиваться в длинные волоски. Из базальных клеток боковых ветвей развиваются ветви неограниченного роста. Кроме того, базальные клетки мутовок ветвей дают начало кортикальным нитям, которые растут вдоль клеток главной оси, совершенно скрывая их, и у некоторых видов, переплетаясь, образуют многослойную обвертку. Из клеток коровых нитей возникают вторичные мутовки боковых ветвей, вставляющиеся между уже имеющимися. На боковых ветвях ограниченного роста формируются половые органы. После оплодотворения из брюшка карпогона вырастают ветвящиеся гонимобласты; конечные клетки их образуют карпоспоры, собранные в тесную группу — цистокарпий. Из карпо-спор развиваются ползучие нити, от которых отходят вертикальные, ветвящиеся нити совсем иного внешнего вида, чем таллом бат-рахоспермума. Эта стадия была описана под родовым названием шантранзия — Chantransia. Она может размножаться посредством моноспор. При благоприятных условиях из верхушечных клеток этого Chantransia-предростка вырастают типичные талломы батра-хоспермума.
Развитие на нитях шантранзии таллома батрахоспермума зависит от продолжительности и интенсивности освещения. Так, при культивировании Batrachospermum moniliforme при фотопериоде менее 14 ч Chantransia-стадия дает только моноспорьг. При освещении ниже 750 Лк наблюдалась только Chantransia-стадия. При длине дня 14 ч и освещении свыше 750 Лк до 4% Chantransia-нитей образовывали гаметофиты, т. е. развивали талломы батрахоспермума. При фотопериоде 15- —16 ч и освещении 750 и 1500 Л к развитие гаметофитов наблюдалось у 40% Chantransia-нитей. У неко-юрых видов Batrachospermum в лабораторных условиях онтогенез от прорастания карпоспор до развития гаметофитов прослежен цитологически. Ввиду мелких размеров ядер содержание ДНК в клетках гаметофитов и Chantransia-нитей, покрашенных по Фёльгену, определялось цитофотометрическим методом. Показано, что Chantransia-стадия диплоидна, а гаметофит гаплоиден. Таким образом, мейоз не мог произойти во время первого деления ядра зиготы, как принималось ранее, а иногда утверждалось и в более современных исследованиях. Можно принять, что мейоз имеет место в переходной области между Chantransia-стадией и гаметофитом. Согласно Мань, Chantransia-стадию батрахоспермума, так же как и базалыгую часть таллома леманеи, следует рассматривать как диплоидное образование, соответствующее тетраспорофиту других красных водорослей.
Другой тип жизненного цикла, хорошо обоснованный,— это тип Bonnemaisonia hamifera, состоящий из последовательности гаметангиальной, карпоспорангиаль-ной и тетраспорангиальной фаз, причем все три морфологически неодинаковы и имеют карпогониальную фазу, развивающуюся на гаметангиальной. Такой тип, иначе гетероморфная смена форм развития, наблюдался у некоторых видов морского рода немалион, отличающегося от леманеи и батрахоспермума многоосевым типом таллома. В слабоветвящихся розовых слизистых талломах немалиона центральная часть занята целым пучком продольных нитей, состоящих из вытянутых бесцветных клеток, от которых радиально расходятся обильно ветвящиеся, богатые хроматофорами нити — ассимиляторы, соединенные слизью мягкой консистенции.
Половые органы, как и у батрахоспермума, развиваются на ассимиляторах. Карпоспорофит и карпоспоры у немалиона, как у леманеи и батрахоспермума, диплоидны. У некоторых видов Nemalion наблюдалась гетероморфная смена генераций: макроскопического гаметофита и микроскопического нитчатого Acrochaetium-подобного тетраспорофита.
Такой же тип развития, в подтверждение гипотезы Мань, описан и для некоторых других представителей немалиальных, обладающих, как и немалион, многоосевыми талломами, но с более компактной структурой и нередко с заметной кальцификацией. Таковы Scinaia furcellata, S. turgida, Helminthora divaricata, Helmintliocladia calvadosii, Chaetangiumfastigiatum. Описанные ранее как два самостоятельных вида рода Acrochaetium A. polyblasum и A. hallandicum оказались соответственно тетраспорофитом и гаметофитом в жизненном цикле одного организма. То же относится и к сильно различающимся морфологически красным водорослям, ранее относимых к разным родам: Falkenbergia rufolanosa и Asparagopsis armata.
Третий тип цикла развития — тип Liagora tetrasporifera1 — складывается из последовательности гаметангиальной и тетраспорангиальной фаз, морфологически неодинаковых, причем тетраспорангиальная фаза развивается на гаметангиальной; мейоз происходит в тетраспорангиях. Такой цикл описан для Helminthocladia agardiana, Н. senegalensis.
Наконец, недавно в культуре Acrochaetium pectinatum впервые для красных водорослей наблюдалось свободное развитие зиготы: содержимое карпогона после оплодотворения выходило из оболочки, прикреплялось к субстрату и прорастало в спорофит. Иногда зигота развивалась в нити гонимобласта с терминальными карпоспорангиями.
Порядок гелидиальные — Gelidiales2
Этот порядок выделил из порядка немалиальных Г. Кюлин на основе наличия тетраспорангиальной фазы, морфологически сходной с гаметангиальной, что позволяло предположить здесь цикл развития типа Polysiphonia. Кюлин придерживался мнения, что, как и у немалиальных, у гелидиума отсутствуют ауксилярные клетки и гонимоблаеты развиваются непосредственно из брюшка оплодотворенного карпогона. Мнения последующих исследователей относительно существования ауксилярных клеток у гелидиума расходятся. Одни утверждают, что поддерживающая клетка, с которой сливается оплодотворенный карпогон, может рассматриваться как ауксилярная, другие же, указывая на то, что это слияние не носит облигатного характера, не признают эту клетку ауксилярной.
Талломы всех гелидиальных имеют одноосевое строение. Все они — обитатели теплых морей. Gelidium cartilagiheum и другие виды этого рода—важные промысловые водоросли: из них в Японии получают кантен.
Порядок криптонемиальные—Cryptonemiales
В отличие от предыдущих порядков здесь представители имеют ауксилярные клетки, которые развиваются до оплодотворения карпогона и обычно рассеяны по таллому на известном расстоянии от карпогона. Связь между ауксилярными клетками и карпогоном осуществляется с помощью более или менее длинных соединительных, спорогенных или ообластемных нитей.
Для представителей порядка криптонемиальных долгое время предполагался жизненный цикл Polysiphonia-типа, при котором гаметангиальная и тетраспорангиальная фазы морфологически идентичны. Такой жизненный цикл был давно описан для Corallina officinalis, тогда как для остальных представителей порядка это предположение не сопровождалось строгими доказательствами, и лишь сравнительно недавно жизненный цикл Callophyllis firma был прослежен в лабораторной культуре и продемонстрировано развитие этой водоросли по Polysiphonia-типу. Гаметангиальные растения несли зрелые карпоспорофиты. Освобождавшиеся карпоспоры прорастали с образованием многослойных базальных дисков, из которых вырастали вертикальные оси и развивались листовидные пластинки, характерные для этого таксона. Приблизительно через три месяца листовидные талломы, возникшие при прорастании карпоспор, продуцировали многочисленные тетраспорангии с крестообразным расположением тетраспор. Из тетраспор, освобождавшихся из тетраспорангиальных растений, в культуре развились гаметангиальные растения со зрелыми карпоспорофитами. Карпоспоры прорастали в новые тетраспорангиальные растения. Гаметангиальная и тетраспорангиальная фазы морфологически идентичны. Исследования других представителей порядка криптонемиальных выявили иной жизненный цикл, при котором гаметангиальная и тетраспорангиальная фазы морфологически различались.
Среди криптонемиальных, как и среди немалиальных, встречаются формы не только одноосевого, но и многоосевого строения.
Порядок гигартинальные—Gigartinales
Порядок характеризуется тем, что ауксилярные клетки дифференцируются до оплодотворения в непосредственной близости от карпогона, так что прокарпий имеется; в качестве ауксилярной функционирует клетка, участвующая в формировании таллома, а не специальная — добавочная. Размножение можно рассмотреть на примере рода анфельция. Зрелые мужские и женские гаметофиты представлены вертикальными, в различной степени разветвленными многоосевыми талломами, вырастающими от хорошо развитых базальных корочек. На мужских талломах в плодущих участках зрелые антеридии образуют один ряд вытянутых клеток. При помещении живого материала в морскую воду под микроскопом можно наблюдать, как вытянутые спермации с силой отбрасываются от таллома на значительное расстояние. На женских талломах плодущие участки находятся вблизи верхушки. Прокарпий Ahnfeltia concinna состоит из крупной поддерживающей клетки и трехклеточной карпогонной ветви, верхняя клетка которой представляет собой карпогон. Поддерживающая клетка функционирует как ауксилярная. После оплодотворения карпогона и слияния его с поддерживающей клеткой, одновременно являющейся ауксилярной, развиваются гонимобласты — карпоспорофиты, врастающие в ткань сердцевины таллома, где они могут сохраняться дольше года до того, как освободятся карпоспоры.
При прорастании карпоспоры образуется корковидный таллом, морфологически отличный от гаметангиальных талломов-гаметофитов в виде вертикальных разветвленных или неразвегвленных пластинок многоосевого строения, растущих от базальных корочек. Приблизительно через три недели после прорастания карпоспор на корковидном талломе начинается формирование тетраспорангиев, в которых развиваются крестообразно расположенные тетраспоры. Тетраспоры прорастают как карпоспоры, образуя сначала корочки, на которых, однако, возникают многоосевые талломы анфельции, сходные с встречающимися в природе. Корочки, развившиеся из тетраспор, никогда не образуют тетраспорангиев, а корочки из карпоспор никогда не дают вертикальных талломов анфельции. Таким образом, у Ahnfeltia concinna гетероморфная смена генераций. У ряда представителей был продемонстрирован жизненный цикл Polysiphonia-типа.
У ряда видов из класса флоридеофициевых было описано аномальное сочетание репродуктивных ортанов; среди этих сообщений многие касаются представителей порядка гигартинальных. Например, у Cystodiniwn purpureum, в норме характеризующегося изоморфной сменой поколений и морфологически сходными гаметофитами и тетраспорофитами, у некоторых растений на одном и том же талломе присутствовали и тстраспорангии, и цистокарпии.
В ряде случаев у одного и того же вида обнаруживаются разные циклы развития. Например, у Gigartina siellata наблюдается или половой цикл с PetraceUis cruenta в качестве тетраспорофита, или же бесполый цикл, по-видимому, с апогамным развитием цистокарпиев.
Порядок родимениальные — Rhodymeniales
Порядок объединяет формы многоосевого строения. Ауксиляр-ные клетки отчленяются от клетки, получившейся в результате деления несущей клетки до оплодотворения карпогона. Ауксилярные клетки — добавочные, специальные клетки, не принимающие участия в построении вегетативного таллома. Есть прокарпии, в состав которых в одних случаях, кроме карпогона, входит одна ауксилярная клетка, в других — две. У одних форм тетраспорангии с тетраэдрическим расположением тетраспор, у других — с крестообразным. Представителем служит род ломен-гария, имеющий трубчатый, полый с перетяжками или без них таллом с боковым ветвлением.
До сравнительно недавнего времени к этому порядку относили широко распространенную по берегам Европы гюдоросль родимению пальчатую {Rhodymenia palmata). Таллом многоосевого типа имеет вид листа, сидящего на суженном основании. В настоящее время эта водоросль под названием пальмария пальчатая выделена в новый порядок пальмари-альные главным образом на основе строения тетраспо-рангиев, с характерными клетками-ножками. Тетраспорангиальный зачаток формируется из коровой клетки таллом