


Основы биохимии
СОДЕРЖАНИЕ
Вопрос 10. Приведите примеры биологически активных пептидов. Какие функции они выполняют?
Вопрос 20. Как связана простетическая группа фосфопротеинов с белковым компонентом. Показать ее на примере аминокислот серина и треонина
Вопрос 30. Энзинопатологии (фенилкетонурия, альбинизм и алкаптонурия)
Вопрос 40. Назвать коферментные формы витаминов биотина и В12, указать в каких обменных процессах они участвуют
Вопрос 46. Напишите формулу сиаловой кислоты. Охарактеризуйте расположением гликопротеинов в биомембране. Какую функцию они выполняют?
Вопрос 50. Написать реакцию НАД-зависимого дгидрирования малата в цикле КРЕБСА
Вопрос 52. Что такое окислительное фосфорелирование? Указать участки сопряжения окисления и фосфорелирования в цепях переноса электронов
Вопрос 64. Напишите фрагменты молекул клетчатки, крахмала. Укажите какой углевод не переваривается в ЖКТ человека и почему?
Вопрос 75. Написать в виде схемы реакции синтеза гликогена
Вопрос 84. Какие вещества необходимы для синтеза жирных кислот на синтаве жирных кислот. Как связан процесс синтеза жирных кислот с обменом углеводов?
Вопрос 88. Какими липопротеинами крови доставляются жиры из печени и кишечника к жировым депо? Каков состав этих липопротеинов?
Вопрос 96. В состав пищи входили пептиды следующего состава: ала-сер-гли-дис -сен-лиз-три-вал-лей. Назовите ферменты и соответствующие связи, которые они расщепляют: а) в желудке, б) в просвете тонкой кишки,
в) в слизистой оболочке тонкой кишки
Вопрос 106. Декарбоксилирование аминокислот. Строение кофермента
декарбоксилаз. Биологическая роль образовавшихся соединений
Вопрос 108. Напишите орнитиновый цикл мочевинообразования. Укажите локализацию процесса
Вопрос 121. Через какие активные соединения клетки гормоны реализуют свое действие?.
Вопрос 126. Охарактеризуйте механизм действия адреналина и глюкагона на липидный обмен
Вопрос 130. Клинико-диагностическое значение определения активности амилазы сыворотки крови и мочи
Вопрос 133. Нарушение прямого и соотношения непрямого билирубина при желтухах. Охарактеризуйте гемолитическую, паренсиматозную и обтурационную желтухи
Вопрос 134. С какими свойствами белков плазмы крови связана их
способность регулировать объем крови и ее онкотическое давление? Какая белковая фракция играет наиболее существенную роль в обеспечении этой функции?
Вопрос 143. Приведите схему строения антител. Обозначьте на схеме участки взаимодействия с антигеном
Список используемой литературы
Вопрос 10. Приведите примеры биологически активных пептидов. Какие функции они выполняют?
Ответ. В последние годы значительно повысился интерес к структуре и функциям встречающихся в свободном состоянии в организме низкомолекулярных пептидов, выполняющих ряд специфических биологических функций. Короткие пептиды, содержащие до 10 аминокислот, принято называть олигопептидами; в то же время полипептиды и белки считаются взаимозаменяемыми, хотя термином «полипептиды» чаще обозначают продукты с мол. м. менее 10000. В некоторых биоактивных пептидах имеются необычные аминокислоты, не встречающиеся в природных белках, или производные обычных аминокислот (гормоны, антибиотики). Мнение о том, что пептиды могут играть роль промежуточных продуктов на пути синтеза белка, не подтвердилось, поскольку этот процесс во всех клетках у всех живых организмов осуществляется de novo матричным путем.
Природные пептиды, наделенные биологической активностью, в зависимости от характера действия и происхождения принято делить на 4 группы: 1) пептиды, обладающие гормональной активностью (вазопрессин, окси-тоцин, кортикотропин, глюкагон, кальцитонин, меланоцитстимулирующий гормон, рилизинг-факторы гипоталамуса и др.; 2) пептиды, принимающие участие в процессе пищеварения (в частности, гастрин и секретин); 3) пептиды, источник которых – α2-глобулиновая фракция сыворотки крови (такие, как ангиотензин, брадикинин и каллидин); 4) нейропептиды.
В последнее время выяснены некоторые закономерности синтеза физиологически активных пептидов из биологически инертных предшественников – белков в результате процесса, называемого посттрансляционной модификацией (постсинтетические превращения белковой молекулы). Известно, например, что ангиотензины (представленные октапептидами), оказывающие выраженное сосудосуживающее действие, образуются из присутствующего в сыворотке крови неактивного белка ангиотензиногена в результате последовательного действия ряда протеолитических ферментов (ренина и особого фермента, участвующего в превращении неактивного ангиотензина I в активный ангиотензин II).
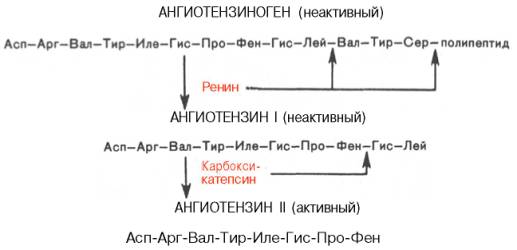
К группе вазоактивных (оказывающих влияние на тонус сосудов) пептидов относятся, кроме того, широко применяемые в медицинской практике брадикинин и каллидин.
Брадикинин представляет собой нонапептид:
H–Apг–Про–Про–Гли–Фен–Сер–Про–Фен–Apг–ОН.
Каллидин представлен декапептидом, образующимся из неактивного плазменного белка кининогена, и отличается от брадикинина присутствием на N-конце еще одного аминокислотного остатка (Лиз):
Н–Лиз–Арг–Про–Про–Гли–Фен–Сер–Про–Фен–Apг–ОН.
Совсем недавно из экстрактов ткани предсердия (но не из желудочков сердца) человека и животных были выделены биологически активные пептиды, регулирующие тонус сосудистой системы и электролитный обмен. Физиологический эффект их оказался противоположным влиянию системы ренин–ангиотензин–альдостерон. Он выражается в сосудорасширяющем действии, усилении клубочковой фильтрации и стимуляции выведения натрия и хлоридов за счет угнетения их реабсорбции в канальцах. Эти пептиды получили название а триопептидов (от лат. atrio – предсердие). Они построены из разного числа аминокислот (от 23 до 100), но обязательным условием для проявления биологического эффекта является наличие в молекуле 17-членной кольцевой структуры, образующейся за счет дисуль-фидной связи между остатками цистеина.
Внутриклеточным посредником действия атриопептидов оказался циклический гуанозинмонофосфат (цГМФ), синтез которого осуществляется в результате активирования мембранного фермента гуанилатциклазы; действие аденилатциклазы, напротив, тормозится под влиянием атриопеп-тидов.
Во всех животных тканях и в некоторых растениях широко распространен низкомолекулярный трипептид глутатион, функции которого пока не выяснены достаточно полно, хотя он открыт сравнительно давно. Глута-тион представляет собой атипичный трипептид (в котором в образовании одной из пептидных связей участвует не α-карбоксильная, а γ-карбок-сильная группа глутамата) следующего строения: γ-глутамил-цистеинил-глицин:

Цистеин является составной частью глутатиона, поэтому последний может находиться в восстановленной (SH) и в окисленной (S-S) формах (сокращенно обозначаются Г-SH и Г-S-S-Г), что, по-видимому, имеет отношение к биологической роли глутатиона в организме.
Интерес к природным пептидам в значительной степени обусловлен необычно высокой их биологической активностью. Они оказывают мощное фармакологическое действие на множество физиологических функций организма. В то же время были замечены низкая стабильность и быстрый распад их в организме при физиологических значениях рН среды. Все это способствовало развитию исследований как в области препаративного выделения природных пептидов из органов и тканей (включая получение биологически активных пептидов из предшественников методами ограниченного протеолиза ряда хорошо известных гормонов), так и в области химического синтеза. Получение ряда биологически активных нейропепти-дов из гормонов гипофиза, в частности эндорфинов и энкефалинов, наделенных мощным обезболивающим действием (путем связывания рецепторов определенных клеток мозга), в сотни и тысячи раз превосходящим аналгезирующий эффект морфина.
Из ткани мозга выделен также δ-пептид сна; ряд других нейропептидов принимает участие в биохимических механизмах памяти, страха, обучения и т.д. Для повышения стабильности пептидов при введении в организм предприняты попытки химического синтеза пептидов, в которых один или несколько аминокислотных остатков L-ряда замещают остатками D-аминокислот. Подобная замена, не вызывая снижения биоактивности, защищает пептид от воздействия протеиназ тканей, способствуя пролонгированию эффекта препарата.
Среди естественно встречающихся небольших пептидов следует указать на антибиотик грамицидин S, выделенный из Bacillus brevis и представляющий собой циклический декапептид:
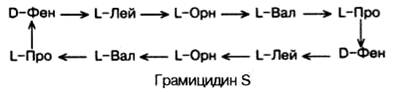
Как видно, в структуре грамицидина S имеются 2 остатка орнитина (Орн), производные аминокислоты аргинина и 2 остатка неприродных D-изомеров фенилаланина. Стрелки указывают направление синтеза от NН2-групп к СООН-группам каждого остатка, и вследствие цикличности грамицидин S не имеет конца.
Широкое применение, особенно в пищевой промышленности, в качестве заменителя сахара получил искусственный (генноинженерный синтез) ди-пептид, состоящий из L-изомеров аспарагиновой кислоты и метилового эфира фенилаланина, названный аспартамом:
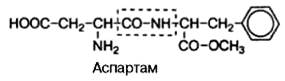
Аспартам в сотни раз слаще сахара и легко распадается в организме на две свободные аминокислоты, абсолютно безвредные для организма; поэтому он рекомендован в качестве заменителя сахара больным диабетом. Это пример пептида, наделенного огромным биологическим эффектом.
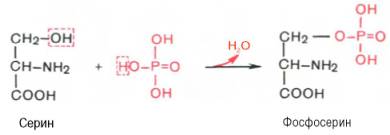
Вопрос 20. Как связана простетическая группа фосфопротеинов с белковым компонентом. Показать ее на примере аминокислот серина и треонина
Ответ. К белкам этого класса относятся казеиноген молока, в котором содержание фосфорной кислоты достигает 1%; вителлин, вителлинин и фосвитин, выделенные из желтка куриного яйца; овальбумин, открытый в белке куриного яйца; ихтулин, содержащийся в икре рыб, и др. Большое количество фосфопротеинов содержится в клетках ЦНС. Фосфопротеины занимают особое положение в биохимии фосфорсодержащих соединений не только в результате своеобразия структурной организации, но и вследствие широкого диапазона функций в метаболизме. Характерной особенностью структуры фосфопротеинов является то, что фосфорная кислота оказывается связанной сложноэфирной связью с белковой молекулой через гидроксильные группы β-оксиаминокислот, главным образом серина и в меньшей степени треонина. На одну молекулу белка обычно приходится 2–4 остатка фосфата.
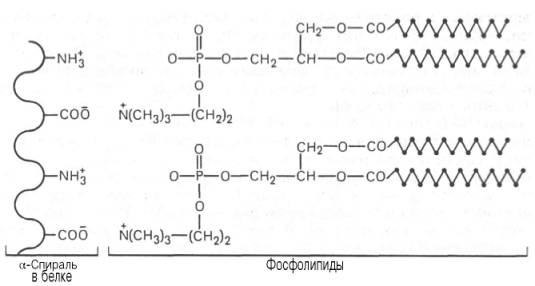
Рис. 1. Ионный тип связи между белками и фосфолипидами.
Новые данные свидетельствуют о том, что в клетках фосфопротеины синтезируются в результате посттрансляционной модификации, подвергаясь фосфорилированию при участии протеинкиназ. Таким образом, уровень фосфопротеинов в клетке зависит в значительной степени от регулирующего действия ферментов, катализирующих фосфорилирование (протеин-киназы) и дефосфорилирование (протеинфосфатазы). Следует отметить, что фосфопротеины содержат органически связанный, лабильный фосфат, абсолютно необходимый для выполнения клеткой ряда биологических функций. Кроме того, они являются ценным источником энергетического и пластического материала в процессе эмбриогенеза и дальнейшего постна-тального роста и развития организма.
Особо следует отметить, что некоторые ключевые ферменты, регулирующие процессы внутриклеточного обмена веществ, также существуют как в фосфорилированной, так и в дефосфорилированной форме. Этим подчеркивается значение фосфорилирования–дефосфорилирования в процессах химической модификации макромолекул, участвующих в интегральных процессах метаболизма.
Вопрос 30. Энзинопатологии (фенилкетонурия, альбинизм и алкаптонурия)
Ответ.Фенилкетонурия, фенилпировиноградная олигофрения, наследственное заболевание из группы ферментопатий, в основе которого лежит аномалия аминокислотного обмена вследствие отсутствия или резкого снижения активности фермента фенилаланингидроксилазы. Описана в 1934 норв. учёным А. Фёллингом (A. Foiling) (болезнь Фёллинга). Частота фенилкетонурии – 1 случай на 10–15 тыс. новорождённых; наследуется по аутосомно-рецессивному типу. При фенилкетонурии фенилаланингидроксилаза сохраняет только около 5% активности, в связи с чем нарушается обмен фенилаланина и вследствие этого – тирозина, триптофана и др., накапливаются промежуточные продукты обмена – фенилэтиламин, фенилпировиноградная кислота и др. и возникает дефицит метаболитов, необходимых для нормального функционирования организма. В частности, вероятная причина умственных расстройств – дефицит медиаторов нервной системы (адреналина, норадреналина, серотонина и др.). Таким образом, при фенилкетонурии возникает комплекс взаимосвязанных метаболических расстройств, состоящий из первичного ферментного нарушения и обусловленных им др. нарушений обмена.
Фенилкетонурия проявляется главным образом выраженной олигофренией (идиотией или имбецильностью). Диагностируется в первые дни жизни ребёнка с помощью экспресс-методов – микробиологических или биохимических. Последние основаны на определении пировиноградной кислоты в моче посредством индикаторов (проба Фёллинга). Лечение сводится главным образом к специальной диете (резкое ограничение продуктов, содержащих фенилаланин).
Альбинизм (albinismus) представляет собой врожденное отсутствие кожного пигмента. Этиология и патогенез изучены недостаточно. Известно, что в результате нарушения синтеза ферментов тирозиназы, дофаоксидазы прекращается образование меланина из тирозина, диоксифенилаланина. О сложности механизмов происхождения альбинизма свидетельствуют нередко сопутствующие ему такие аномалии, как врожденная глухота, дефекты интеллекта, патология глаз и другие.
Выделяют тотальный, неполный и частичный альбинизм.
Тотальный альбинизм наследуется аутосомно-рецессивно со средней частотой 1: 10000-20000. Предполагают, что носители мутантного гена составляют 1,5% среди всех нормально пигментированных людей.
Депигментация кожи и придатков наблюдается с рождения, сопровождается сухостью кожи, нарушением потоотделения, иногда гипо- или гипертрихозом, особенно на открытых участках. У больных легко возникают солнечные ожоги, актинический хейлит. Они предрасположены к развитию кератом, эпителиом, телеангиэктазий. Из-за отсутствия пигмента в тканях глаза зрачки кажутся красными. Характерными являются горизонтальный нистагм и выраженная светобоязнь. Часто наблюдаются сходящееся косоглазие, снижение остроты зрения в результате нарушений рефракции, катаракты, возможна микрофтальмия. Нередко наблюдаются бесплодие, иммунодефицит (отсюда частые инфекции), пороки развития, сокращение продолжительности жизни, олигофрения.
Неполный альбинизм (син.: альбиноидизм), в отличие от предыдущей формы, наследуется аутосомно-доминантно, в некоторых случаях — рецессивно. Имеет место снижение активности тирозиназы, но не блокада ее синтеза. Наблюдается гипопигментация кожи, волос, радужки, иногда фотофобия. Других дефектов и аномалий не регистрируется.
Частичный альбинизм (син.: пиебалдизм) наследуется аутосомно-доминантно. Проявления выявляются при рождении. Характеризуется появлением участков ахромии на коже живота, лица, нижних конечностей, прядей седых волос. Депигментированные пятна неправильной формы с резкими границами, на их поверхности имеются мелкие темно-коричневые пятнышки. Вокруг ахромичных пятен кожа может быть пигментированной. Поражений других органов обычно не бывает. Частичный альбинизм является одним из проявлений синдромов Чедиака-Хигаси, Клейна-Ваарденбурга, Титце, Менде, Хермански-Пудлака, Кросса-МакКьюзика-Брина.
Дифференциальный диагноз альбинизма проводят с витилиго, синдромами Клейна-Ваарденбурга, Алеззандрини, Фогта-Коянаги-Харады.
Алкаптонурия (alcaptomiria; алкаптон (гомогентизиновая кислота) + греч. uron моч ) - наследственная болезнь, обусловленная нарушением обмена тирозина вследствие пониженной активности фермента гомогентизиназы и накоплением в тканях организма гомотентизиновой кислоты; проявляется у взрослых пигментацией различных тканей, развитием артрозов, а у детей - лишь темным окрашиванием мочи и иногда ушной серы; наследуется по аутосомно-рецессивному типу.
Вопрос 40. Назвать коферментные формы витаминов биотина и В12, указать в каких обменных процессах они участвуют
Ответ.Биологическая роль. Биотин подробно изучен благодаря работам Ф. Линена. Известные к настоящему времени биотиновые ферменты (т.е. ферменты, содержащие в качестве кофермента биотин) катализируют два типа реакций:
1) реакции карбоксилирования (с участием СО2 или НСО3–), сопряженные с распадом АТФ
RH + HC03– + ATO<=> R-COOH +AflO+H3P04;
2) реакции транскарбоксилирования (протекающие без участия АТФ), при которых субстраты обмениваются карбоксильной группой
R1-COOH + R2H <=> R1H + R2-COOH.
Получены доказательства двустадийного механизма этих реакций с образованием промежуточного комплекса (карбоксибиотинилфермент).
К реакциям первого типа относятся, например, ацетил-КоА- и пируват-карбоксилазные реакции:
C H 3– C O – S - K o A + CO2+ АТФ <=> H O O C – C H 2– C O – K o A + АДФ + Pi.
Пируваткарбоксилаза является высокоспецифичным ферментом, катализирующим уникальную реакцию усвоения СО2 в организме животных. Сущность реакции сводится к пополнению запасов оксалоацетата (щаве-левоуксусная кислота) в лимоннокислом цикле (так называемые «анаплеро-тические», «пополняющие» реакции), т.е. его синтезу из СО2 и пирувата:
Пируват + CO2+ АТФ + H2O —> Оксалоацетат + АДФ + Pi+ 2H+
Реакция протекает в две стадии: на первой стадии, связанной с затратой энергии, СО2 подвергается активированию, т.е. ковалентному связыванию с биотином в активном центре фермента (Е-биотин):
![]()
На второй стадии СО2 из комплекса переносится на пируват с образованием оксалоацетата и освобождением фермента:
![]()
Примером второго типа реакций является метилмалонил-оксалоаце-тат-транскарбоксилазная реакция, катализирующая обратимое превращение пировиноградной и щавелевоуксусной кислот:
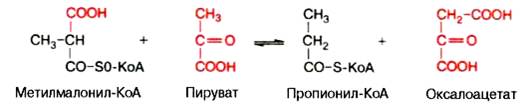
Реакции карбоксилирования и транскарбоксилирования имеют важное значение в организме при синтезе высших жирных кислот, белков, пури-новых нуклеотидов (соответственно нуклеиновых кислот) и др.
Распространение в природе и суточная потребность. Биотин содержится почти во всех продуктах животного и растительного происхождения, главным образом в связанной форме. Богаты этим витамином печень, почки, молоко, желток яйца. В растительных продуктах (картофель, лук, томат, шпинат) биотин находится как в свободном, гак и в связанном состоянии. Для человека и животных важным источником является биотин, синтезируемый микрофлорой кишечника. Суточная потребность взрослого человека в биотине приблизительно 0,25 мг.
Биологическая роль. Выявлены ферментные системы, в составе которых в качестве простетической группы участвуют не свободный витамин В12, а так называемые В12-коферменты, или кобамидные коферменты. Последние отличаются тем, что содержат 2 типа лигандов: метильную группу и 5'-дезоксиаденозин. Соответственно различают метилкобаламин СН3-В12 и дезоксиаденозилкобаламин. Превращение свободного витамина В12 в В12-коферменты, протекающее в несколько этапов, осуществляется в организме при участии специфических ферментов в присутствии в качестве кофакторов ФАД, восстановленного НАД, АТФ и глутатиона. В частности, при образовании 5-дезоксикобаламинового кофермента АТФ подвергается необычному распаду с отщеплением три-фосфатного остатка по аналогии еще с одной единственной реакцией синтеза 5-аденозилметионина из метионина и АТФ. Впервые В12-коферменты были выделены Г. Баркером и сотр. в 1958 г. из микроорганизмов, позже было доказано их существование в тканях животных.
Химические реакции, в которых витамин В12 принимает участие как кофермент, условно делят на 2 группы в соответствии с его химической природой. К первой группе относятся реакции трансметилирования, в которых метилкобаламин выполняет роль промежуточного переносчика метильной группы (реакции синтеза метионина и ацетата).
Синтез метионина требует, помимо гомоцистеина, наличия N5-метил-ТГФК и восстановленного ФАД и протекает согласно уравнению:

Фермент, катализирующий эту реакцию, был открыт в печени человека и ряда животных, а также у микроорганизмов. Получены доказательства, что механизм реакции включает перенос метильной группы N5-СН3-ТГФК на активный центр фермента с образованием метил-В12-фермента и последующий перенос этой группы на гомоцистеин. Блокирование этой реакции, наблюдаемое при авитаминозе В12, приводит к накоплению N5-СН3-ТГФК и соответственно выключению из сферы химических реакций еще одного важного кофермента.
Вторая группа реакций при участии В12-коферментов заключается во внутримолекулярном переносе водорода в реакциях изомеризации. Механизм этих реакций соответствует схеме:

Видно, что протон водорода движется (перемещается) между двумя соседними атомами углерода и не обменивается с протонами воды. Предполагают, что сначала водород от субстрата переносится на 5-дезокси-кобаламин, а затем обратно на субстрат, меняя местоположение. Например, глутаматмутазная реакция (взаимопревращения глутаминовой и β-метиласпарагиновой кислот), метилмалонилмутазная реакция (обратимое превращение метилмалонил-КоА в сукцинил-КоА), глицерол- и диол-дегидратазные реакции, ферментативные реакции восстановления рибо-нуклеотидов до дезоксирибонуклеотидов и др. В организме человека из указанных процессов открыта только реакция изомеризации метил-малонил-КоА в сукцинил-КоА.
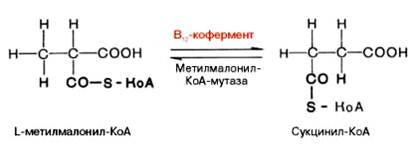
Следует подчеркнуть, что реакция изомеризации метилмалонил-КоА требует наличия 5'-дезоксиаденозилкобаламина в качестве кофермента, в то время как реакция метилирования нуждается в метилкобала-мине. Этими обстоятельствами могут быть объяснены некоторые биохимические симптомы недостаточности витамина В12, в частности метил-малонилацидурия и гомоцистинурия. Кроме того, описаны болезни, обусловленные наследственными дефектами синтеза только дезоксиаденозил-кобаламина или обоих В12-коферментов; в этих случаях даже 1000-кратная доза витамина В12 не оказывала лечебного эффекта. В настоящее время высказывается предположение о более широком участии В12-коферментов в ферментативных реакциях трансметилирования, дезаминирования (например, этаноламиддезаминазная реакция) и др. Предстоит, однако, приложить немало усилий, чтобы выяснить молекулярные механизмы действия витамина В12 на процесс кроветворения. Положительный эффект при лечении пернициозной анемии полусырой печенью обусловлен, как стало известно, наличием витамина В12, хотя следует указать, что большего лечебного эффекта можно добиться при одновременном введении внутреннего фактора слизистой оболочки желудка.
Распространение в природе и суточная потребность. Витамин В12 является единственным витамином, синтез которого осуществляется исключительно микроорганизмами; ни растения, ни ткани животных этой способностью не наделены. Основные источники витамина В12 для человека – мясо, говяжья печень, почки, рыба, молоко, яйца. Главным местом накопления витамина В12 в организме человека является печень, в которой содержится до нескольких миллиграммов витамина. В печень он поступает с животной пищей, в частности с мясом, или синтезируется микрофлорой кишечника при условии доставки с пищей кобальта. Суточная потребность в витамине В12 для взрослого человека составляет около 3 мкг (0,003 мг).
Вопрос 46. Напишите формулу сиаловой кислоты. Охарактеризуйте расположением гликопротеинов в биомембране. Какую функцию они выполняют?
Ответ.Гликопротеины – сложные белки, содержащие, помимо простого белка или пептида, группу гетероолигосахаридов. В настоящее время их принято называть гликоконъюгатами. В состав гликоконъюгата входит углеводный компонент (гликановая фракция), ковалентно связанный с неуглеводной частью (агликановая фракция), представленной белком, пептидом, аминокислотой или липидом.
Повышенный интерес к науке об углеводах – гликобиологии – в настоящее время объясняется открытием существенной роли изменений структуры гликоконъюгатов в развитии таких болезней, как рак, иммунодефицит человека, ревматоидные артриты, астма и др. Оказалось, что нарушение реакции гликозилирования двух главных классов глико-конъюгатов (гликопротеинов и ганглиозидов) приводит или к накоплению предшественников этих веществ, или к синтезу «укороченных» сахарных цепей гликоконъюгатов. Более того, установлено, что во взаимодействии между некоторыми вирусами и клетками-мишенями главную роль играют углеводные компоненты. В частности, гликопротеин gp120 вируса иммунодефицита человека (содержит большой процент углевода) имеет высокое сродство к гликопротеину CD4Т-лимфоцита. В этом взаимодействии, узнавании, являющемся высокоспецифичным, гликозилированные фрагменты, вероятнее всего, играют важную патогенетическую роль. Известно также, что при ревматоидных артритах часто синтезируются аномальные антитела (аномальные иммуноглобулины – все гликопротеины) с необычайно короткими сахарными цепями, что вызывает стимуляцию иммунной системы против самого организма. Из этих примеров видно, что, помимо гликобиологии, наступило время признания и таких наук, как гликопа-тология и гликотерапия.
Помимо гликопротеинов, различают также протеогликаны, состоящие из белка и гликозаминогликанов (прежнее название мукополиса-хариды); последние состоят из цепей сложных углеводов: аминосахаров, уроновых кислот, серной кислоты и отдельных моносахаридов. Типичными гликозаминогликанами являются гиалуроновая кислота, хондроитинсерная кислота и гепарин.
К типичным гликопротеинам относят большинство белковых гормонов, секретируемые в жидкие среды организма вещества, мембранные сложные белки, все антитела (иммуноглобулины), белки плазмы крови, молока, овальбумин, интерфероны, факторы комплемента, группы крови, рецеп-торные белки и др. Из этого далеко не полного перечня гликопротеинов видно, что все они выполняют специфические функции: обеспечивают клеточную адгезию, молекулярное и клеточное узнавание, антигенную активность опухолевых клеток, оказывают защитное и гормональное, а также антивирусное действие.
Химический состав гликопротеинов более или менее установлен, структура определена только у ряда из них. К полипептиду присоединяются гетероолигосахаридные цепи, содержащие от 2 до 10, реже 15 мономерных остатков гексоз (галактоза и манноза, реже глюкоза), пентоз (ксилоза, арабиноза) и конечный углевод, чаще всего представленный N-ацетилга-лактозамином, L-фукозой или сиаловой кислотой; в отличие от про-теогликанов гликопротеины не содержат уроновых кислот и серной кислоты.
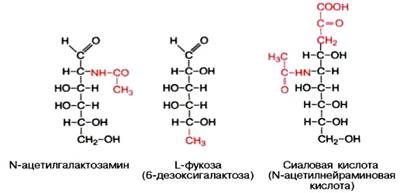
Типы связей между углеводными компонентами и белками определены только у ряда гликопротеинов, аминокислотный состав и структура которых известны (иммуноглобулины, гормоны); они включают О-гликозидные связи (с ОН-группами серина, треонина и оксилизина), N-гликозидные связи (с амидными группами аспарагина, реже глутамина или ω-NH2-группами лизина и аргинина) и эфирные гликозидные связи со свободными СООН-группами глутаминовой и аспарагиновой кислот.
Синтез гликопротеинов осуществляется в рибосомах эндоплазматиче-ского ретикулума (в цистронах), затем присоединяются сахарные цепи (постсинтетическое гликозилирование), и далее белок транспортируется до биомембран клетки и включается в состав мембранных белков или секре-тируется.
Углеводные компоненты соединены ковалентно с азотом аспарагина молекулы белка. Однако предварительно олигосахаридная часть соединяется с липидным переносчиком – долихолфосфатом (липид, содержащий от 15 до 20 изопреновых остатков) и переносится на полипептидную цепь в эндоплазматическом ретикулуме, при этом транспортер освобождается:

Долихолфосфат (n = 15-30)
Синтезированные гликопротеины далее переносятся в аппарат Гольджи, где осуществляются окончательное гликозилирование и сортировка по назначению.
Структура одного из нескольких гетероолигосахаридных остатков в молекуле гликопротеинов, в частности иммуноглобулинов, может быть представлена в виде следующей схемы (использованы сокращения: Глю – глюкоза, NАцГлюА – N-ацетилглюкозамин; Гал – галактоза; Ман – манно-за; NАцНейр – N-ацетилнейраминовая кислота):
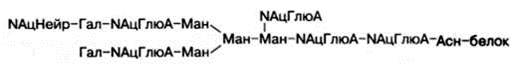
Рассмотрим известные к настоящему времени данные о синтезе, строении (структуре) и свойствах ряда гликопротеинов.
Интерфероны. Интерфероны – это ингибиторы размножения многих типов вирусов. Открыто несколько типов интерферонов (α, β и γ), некоторые из них получены методами генетической инженерии. Это сравнительно небольшие сложные белки с мол. массой у разных видов животных и человека от 25000 до 38000–40000). Они образуются в клетке в ответ на внедрение вирусной нуклеиновой кислоты, ограничивая вирусную агрессию (инфекцию). Известно также, что группа видоспецифических α-интерфе-ронов синтезируется макрофагами, в то время как γ-интерферон продуцируется Т-клетками и стимулируется интерлейкином-2. Показано также, что γ-интерферон в свою очередь повышает цитотоксическую активность макрофагов, Т-клеток и естественных клеток-киллеров. Интерфероны наделены антипролиферативной активностью и считаются основными защитными белками не только против вирусной инфекции, но и при опухолевых поражениях.
Следует отметить, однако, что до сих пор не раскрыты молекулярные механизмы, при помощи которых интерфероны тормозят размножение вирусов. Известно только, что интерфероны ингибируют биосинтез всех белков (и хозяйских, и вирусных), вероятнее всего, на уровне процесса трансляции. Возможно, что интерферон индуцирует синтез особого белка-ингибитора, который затем связывается с рибосомами и блокирует трансляцию, или интерферон переводит один из активных эукариотических белковых факторов инициации в неактивный фактор путем фосфорилиро-вания.
Иммуноглобулины. Иммуноглобулины, или антитела, также относятся к классу гликопротеинов, выполняют защитную функцию, обезвреживая поступающие в организм чужеродные вещества – антигены любой химической природы. Синтезируются иммуноглобулины плазматическими клетками, образовавшимися из лимфоцитов. Учение об иммунитете оформилось в самостоятельную науку – иммунологию, изучающую структуру и функции антител вообще и иммуноглобулинов в частности. Мы представим современные сведения о некоторых физико-химических свойствах и структуре иммуноглобулинов человека (табл. 2.4). Различают 5 классов иммуноглобулинов: IgG, IgM, IgA, IgD и IgE. Детально изучены структура и функция IgG.
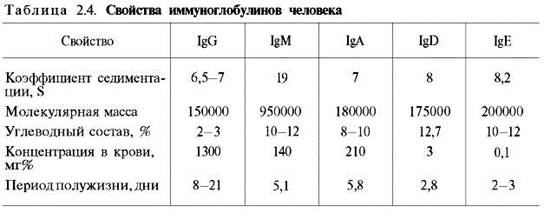
Разные классы иммуноглобулинов сильно различаются не только по молекулярной массе, но и по концентрации в крови; имеются данные, что различаются они и по биологическим свойствам.
Подробно изучена структура IgG. Он имеет Y-образную форму и тет-рамерное строение; состоит из двух идентичных легких L-цепей (от англ. light) и двух идентичных тяжелых Н-цепей (от англ. heavy) с мол. массой 23000–24000 и 50000–70000 соответственно. Известно также, что каждая из этих цепей имеет 2 типа доменов – вариабельные (V) участки, состоящие из 108 аминокислотных остатков, и константные (С) участки, состоящие из 110 и 350 аминокислотных остатков соответственно в L- и Н-цепях (рис. 2).
Из других гликопротеинов, выполняющих ряд важнейших биологических функций, следует отметить все белки плазмы крови (за исключением альбуминов), трансферрин, церулоплазмин, гонадотропный и фолликуло-стимулирующие гормоны, некоторые ферменты, а также гликопротеины в составе слюны (муцин), хрящевой и костной тканей и яичного белка (овомукоид). Углеводные компоненты, помимо информативной функции, значительно повышают стабильность молекул, в состав которых они входят, к различного рода химическим, физическим воздействиям и предохраняют их от действия протеиназ, определяя тем самым биологическую роль гликопротеинов. Являясь составной частью клеточной мембраны, гликопротеины участвуют, кроме того, в иммунологических реакциях, ионном обмене, процессах межклеточной адгезии и т.д.
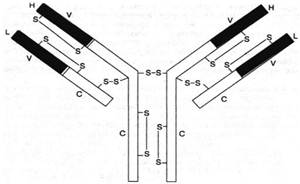
Рис. 2. Структура IgG человека. Показаны легкие (L) и тяжелые (Н) цепи, дисульфидные связи и вариабельные V (красные) и константные С (светлые) участки.
Вопрос 50. Написать реакцию НАД-зависимого дгидрирования малата в цикле КРЕБСА
Ответ. Цикл трикарбоновых кислот впервые был открыт английским биохимиком Г. Кребсом. Он первым постулировал значение данного цикла для полного сгорания пирувата, главным источником которого является гликолитическое превращение углеводов. В дальнейшем было показано, что цикл трикарбо-новых кислот является тем центром, в котором сходятся практически все метаболические пути. Таким образом, цикл Кребса – общий конечный путь окисления ацетильных групп (в виде ацетил-КоА), в которые превращается в процессе катаболизма большая часть органических молекул, играющих роль «клеточного топлива»: углеводов, жирных кислот и аминокислот.
Образовавшийся в результате окислительного декарбоксилирования пирувата в митохондриях ацетил-КоА вступает в цикл Кребса. Данный цикл происходит в матриксе митохондрий и состоит из восьми последовательных реакций (рис. 3). Начинается цикл с присоединения ацетил-КоА к оксалоацетату и образования лимонной кислоты (цитрата). Затем лимонная кислота (шестиуглеродное соединение) путем ряда дегидрирований (отнятие водорода) и двух декарбоксилирований (отщепление СО2) теряет два углеродных атома и снова в цикле Кребса превращается в оксалоацетат (четырехуглеродное соединение), т.е. в результате полного оборота цикла одна молекула ацетил-КоА сгорает до СО2 и Н2О, а молекула окса-лоацетата регенерируется. Рассмотрим все восемь последовательных реакций (этапов) цикла Кребса.
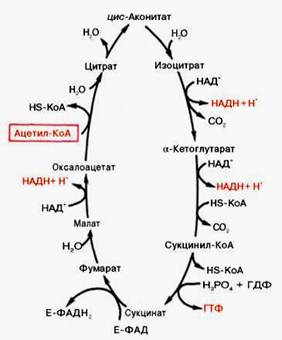
Рис. 3. Цикл трикарбоновых кислот (цикл Кребса).
Первая реакция катализируется ферментом цит-рат-синтазой, при этом ацетильная группа ацетил-КоА конденсируется с оксалоацетатом, в результате чего образуется лимонная кислота:
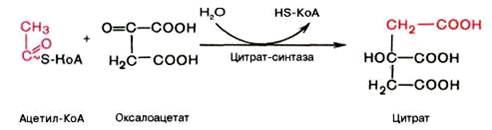
По-видимому, в данной реакции в качестве промежуточного продукта образуется связанный с ферментом цитрил-КоА. Затем последний самопроизвольно и необратимо гидролизуется с образованием цитрата и HS-KoA.
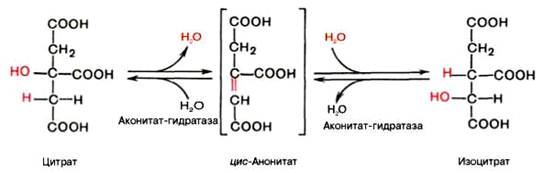
В результате второй реакции образовавшаяся лимонная кислота подвергается дегидратированию с образованием цис-аконитовой кислоты, которая, присоединяя молекулу воды, переходит в изолимонную кислоту (изоцитрат). Катализирует эти обратимые реакции гидратации–дегидратации фермент аконитатгидратаза (аконитаза). В результате происходит взаимоперемещение Н и ОН в молекуле цитрата:
Третья реакция, по-видимому, лимитирует скорость цикла Кребса. Изолимонная кислота дегидрируется в присутствии НАД-зависимой изо-цитратдегидрогеназы.
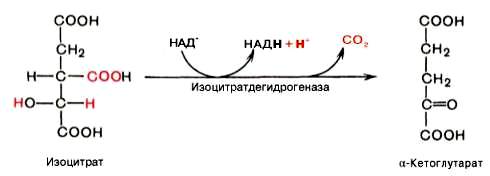
В ходе изоцитратдегидрогеназной реакции изолимонная кислота одновременно декарбоксилируется. НАД-зависимая изоцитратдегидрогеназа является аллостерическим ферментом, которому в качестве специфического активатора необходим АДФ. Кроме того, фермент для проявления своей активности нуждается в ионах Mg2+или Мn2+.
Во время четвертой