


Особенности биологии и экологии Аphroditiformia Баренцева моря
До недавних пор про подкласс Чешуйчатые Многощетинковые черви (Aphroditiformia), или Афродита, она же «морская мышь» была известна лишь систематика, анатомические и географические особенности, связанные с местом обитания; в настоящее же время к «морской красавице» возник неподдельный интерес, связанный с удивительными особенностями этого червячка. Наиболее значимой она оказалась в области не только рыбного промысла, в качестве деликатесов пищи(пескожил Arenicola marina), но и в научной деятельности, а точнее в фотонной инженерии. Афродита обладает более сложной структурной организацией, по сравнению с предшествующими типами,
Цель: Изучить биологию и экологию чешуйчатых многощетинковых червей рода полинойд.
Задачи:
1)Рассмотреть систематическое положение Чешуйчатых червей;
2) Рассмотреть морфо-функциональные особенности червей данного рода;
3) Описать эволюцию Aphroditiformia.
1. ПЕРВЫЕ ТРАКТОВКИ СИСТЕМАТИЧЕСКОГО ПОЛОЖЕНИЯ APHRODITIFORMIA
Чешуйчатые многощетинковые черви с давних пор привлекают внимание исследователей. Уже Линней в десятом издании своей Systema naturae (1758) выделил в качестве самостоятельного рода Aphrodita aculeata, который считается типовым для всей группы чешуйчатых червей.
Афродита, или так называемая морская мышь, имеет продолговатую форму тела, нередко превышающую в длину 10 см, и со спины густо прикрыта войлоком из длинных капиллярных щетинок. Этот червь обычно почти полностью закапывается передним концом тела в мягкий ил, так что на его поверхности виден лишь небольшой пучок удлиненных спинных усиков нескольких задних сегментов. Он невольно поражает своей изумительной и неподдельной красотой. Особое впечатление производят его длинные волосовидные щетинки, образующие густую бахрому по бокам тела. Анатомию Aphrodita aculeata впервые изучил академик П. Паллас, заведывавший зоологическими коллекциями Кунсткамеры Петербургской Академии наук. В своих Miscellanea Zoologica (1766) он обоснованно предложил объединить Aphrodita с другими многощетинковыми червями — Nereis и Serpula — в особую самостоятельную группу, отделив ее от Mollusca, с которыми до этого они рассматривались совместно.
Начиная с самых первых классификаций аннелид многощетинковые черви, несущие на спинной стороне тела плоские чешуйки, или элитры, всегда выделялись как отдельное семейство или ряд семейств, а в некоторых случаях и как самостоятельный отряд. Наиболее обстоятельная первая характеристика чешуйчатых червей дана в работе французских натуралистов Одуэна и Мильн-Эдвардса, хотя их семейство Aphroditiens насчитывало в тот период еще крайне небольшое количество известных видов и родов. В дальнейшем сведения по группе чешуйчатых начали быстро расширяться, и они некоторыми учеными принимались за самостоятельный отряд Aphroditea или Aphroditacea, Весьма высокий таксономический ранг чешуйчатым червям придал и Катрфаж, который семейства Aphroditiens и Palyriens выделил в особый подотряд Erraticae aberrantes, противопоставив его всем остальным эррантным полихетам (подотряд Erraticae propriae). Это было сделано им на том основании, что у чешуйчатых червей наблюдается неоднородность сегментов (anneaux dissimilaris), выражающаяся в чередовании усиковых и чешуйчатых сегментов, в то время как у всех остальных эррантных полихет все сегменты более или менее однотипные (anneaux similaires ou subsimilaires).
Предложены и некоторые другие трактовки систематического положения чешуйчатых червей. Так, наиболее крупный немецкий полихетолог Элерс считал чешуйчатых червей лишь отдельным семейством в составе весьма обширного отряда Nereidea, объединяющего всех современных эррантных полихет. При этом указанный автор для семейства использует то же название Aphroditea, которое ранее Кинбергом было предложено для обозначения отряда. Французский исследователь Клапаред также рассматривал чешуйчатых червей лишь как семейство — Aphroditiens, но в его составе выделял 6 триб — Aphrodites, Polynoides, Pholoides, Acoetides, Sigalionides и Polylepides. Шведский исследователь Мальмгрен различал 4 самостоятельных семейства чешуйчатых червей — Aphroditidae, Polynoidae, Acoetidae (=Polyodontidae) и Sigalionidae, которые в дальнейшем некоторыми авторами понимались как подсемейства. Как видно из всего сказанного, единое мнение о классификации чешуйчатых червей отсутствует. Подробная синонимия всех предложенных в прошлом столетии названий различных таксономических категорий дана в работе Дарбу. В более позднее время американскими учеными все чешуйчатые черви были выделены в надсемейство Aphroditoidea, однако это же название иногда употребляется и в более широком понимании, а именно для объединения почти всех эррантных полихет, за исключением глицерид и эвяицид.
В данной работе для чешуйчатых червей я использую весьма удачный термин Aphroditiformia, предложенный Левинсеном. По этому каталогу подотряд Aphroditiformia состоит из трех надсемейств: Aphroditacea (с 6 семействами), Chrysopetalacea (с 2 семействами) и Pisionacea (с 1 семейством). Общая схема классификации выглядит следующим образом (после названия семейств в скобках первая цифра обозначает количество родов вторая — видов):
Подотряд APHRODITIFORMIA Надсем. Aphroditacea
Сем. Aphroditidae (7, 63)
Сем. Polynoidae (93, 539)
Сем. Sigalionidae (17, 156)
Сем. Pholoididae (1, 4)
Сем. Eulepethidae (4, 12)
Сем. Polyodontidae (6, 39)
Надсем. Chrysopetalacea
Сем. Chrysopetalidae (4, 24)
Сем. Palmyridae (1,2)
Надсем. Pisionacea Сем. Pisionidae (4, 14)
Надсемейство Aphroditacea является центральной группой в филогенетическом отношении, обладает большим количеством родов и видов, отличающихся морфологическим разнообразием. Наибольшее в этом надсемействе значение имеет семейство Polynoidae. Надсемейство Chrysopetalacea представляет собой как бы связующее звено с высокоспециализированным семейством Amphinomida. Надсемейство Pisionacea занимает промежуточное положение между Aphroditiformia и Glyceriformia,причем по личиночному развитию пизиониды ближе всего к чешуйчатым червям.
2. КРАТКИЙ МОРФО-ФУНКЦИОНАЛЬНЫЙ ОЧЕРК APHRODITIFORMIA
2.1 ФОРМА ТЕЛА И РАЗМЕРЫ
Тело чешуйчатых червей, как правило, заметно сплющено дорсовентрально. Спинная сторона, прикрытая спинными чешуйками, или элитрами, несколько выпуклая (сводчатая), а брюшная — более плоская, с медиальным продольным желобком. К переднему и особенно к заднему концам тело немного суживается; на заднем конце тела все сегменты становятся более короткими и узкими. Многие чешуйчатые черви обладают относительно коротким телом, имеют овальную форму; у других же, наоборот, оно сильно вытянутое («червеобразное»). Форма тела связана с образом жизни. Удлиненное тело характерно для червей, глубоко зарывающихся в грунт (многие Sigalionidae), строящих собственные длинные трубки (Polyodontidae), а также для червей, ведущих комменсальный образ жизни (некоторые Polynoidae). У чешуйчатых червей наблюдается вполне определенная тенденция к стабилизации количества сегментов и к укорочению тела, что для них, бесспорно, является эволюционно прогрессивным моментом.
Форма тела чешуйчатых червей, в противоположность многим другим поли-хетам, весьма постоянная, не подверженная сколько-нибудь заметному удлинению и сжиманию при передвижениях, причем она мало изменяется и при фиксации. Некоторые полиноиды с укороченным телом (в основном Lepidonotus) при фиксации сворачиваются в колечко (уже по этой особенности они хорошо распознаются среди прочих полиноид).
Размеры чешуйчатых червей варьируют. Представители семейства Aphroditidae s. str. характеризуются довольно крупными размерами. Так, Aphrodita aculeata, весьма обычная у берегов Европы, нередко имеет в длину около 100 мм, при ширине 40—50 мм (отмечены экземпляры до 180 мм длины), a Laetmonice producta из южного полушария достигает длины 135 мм и ширины до 40— 65 мм, включая параподии со щетинками. В семействе Polynoidae также наблюдаются гиганты. В частности, к ним относится калифорнийская полиноида Hololepida magna длиной 250 мм. Из антарктических районов в коллекциях ЗИН АН СССР имеются экземпляры Eulagisca gigantea, достигающие в длину 210 мм при ширине 60 мм без щетинок и 100 мм со щетинками. В арктических морях одна из наиболее крупных полиноид — Eunoe nodosa — до 100 мм длины и 35 мм ширины. Средняя же длина большинства видов полиноид около 30—40 мм. Наиболее тонкие и длинные полиноиды — Lepidasthenia (до 200 мм длины), характеризующиеся также наибольшим количеством сегментов.
Великанами среди полихет справедливо считаются некоторые Polyodontidae, живущие в очень крупных длинных илистых трубках, в связи, с чем их иногда называют «илистыми сосисками»; так, имеются указания, что они достигают в длину 2 метров при ширине 2 см. Среди сигалионид немало видов до 300 мм длины при ширине 8—9 мм (Sigalion squamatum и др.). Но наряду с такими крупными формами среди чешуйчатых червей есть виды, которые характеризуются крайне небольшими размерами., не превышающими 20—25 мм. При этом длина тела не коррелирует с количеством сегментов. Например, Pholoe minuta с 70 сегментами имеет в длину максимум 20 мм, а у упомянутого выше Laetmonice producta, достигающего 135 мм, количество сегментов не свыше 50; у Eulagisca gigantea и Harmothoe Imbricata количество сегментов не превышает 37—38, в то же время длина особей первого вида иногда превосходит 200 мм, а второго — достигает только 50 мм, обычно около 30 мм.
Наиболее разнообразна окраска элитр. Последние могут иметь пестрый или узорчатый рисунок, непосредственно связанный со скульптурой самих элитр. Исключительно красив бросающийся в глаза рисунок в виде продольных полос на элитрах Hyperhalosydna striata. Уже по этому рисунку удается безошибочно определить вид. Иногда элитры бесцветные и полупрозрачные, — например, у видов Aphrodita, элитры которых прикрыты наружным войлоком из волосовидных щетинок, или у Bylgides sarsi, у которого наличие тонких и нежных элитр связано с его факультативным планктонным образом жизни. У Psammolyce, зарывающегося в песок, элитры, как правило, прочно обклеены мелкими песчинками. Это, по всей видимости, своего рода маскировка, (предохраняющая червей от врагов). Благодаря относительно темной окраске тела, в особенности элитр, соответствующей общему фону субстрата, на котором поселяются полиноиды, последних не всегда удается быстро обнаружить. К этому следует добавить, что элитры крупных ползающих по поверхности грунта полиноид часто покрыты разнообразными эпифитами и различными комменсалами, что еще больше скрывает их на морском дне.
Щетинки большей частью золотистого или светло-янтарного оттенка, могут быть и совершенно прозрачными. Наряду с этим крупные спинные игловидные щетинки Aphrodita почти черного цвета.
2.2 СЕГМЕНТАЦИЯ И ДЕЛЕНИЕ ТЕЛА НА ОТДЕЛЫ
Тело многощетинковых червей поделено на сегменты (членики), которые внешне хорошо распознаются по наличию на каждом из них по одной паре параподий, снабженных обычно различными придатками. Наружная кольчатость тела тесно связана с внутренними септами (диссепиментами),: обособляющими друг от друга отдельные сегменты (метамеры, или сомиты) (рис. 1). В каждом сегменте, как правило, закономерно повторяются наборы внутренних органов; комплекс нервных образований одного сегмента обозначается как невросомит, сплетение кровеносных сосудов — ангиосомит, система мышечных волокон — миосомит и т. д.; большей частью метамерно располагаются также выделительные органы и гонады. У чешуйчатых червей кишечник в каждом сегменте имеет боковые слепые выросты (карманы, или дивертикулы), так что и он у них обладает вполне определенной метамерией (рис. 2).
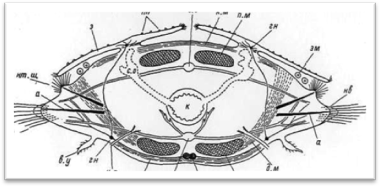
Рис. 1. Поперечный разрез через туловищный сомит (чешуйчатый сегмент).
Таким образом, наружная кольчатость, или наружная метамерия, червей обычно соответствует внутренней метамерии.1
Полная гомономность (однородность) всех туловищных сегментов у многощетинковых червей никогда не наблюдается. Наибольшая гомономность сегментов характерна для бродячих (эррантных) полихет, но и у них она в той или другой степени нарушена. Следует различать гетерономность первичную и вторичную. Первичная гетерономность возникает еще в процессе метаморфоза личинки и связана с образованием ларвальных и постларвальных сегментов, имеющих разное происхождение. Ларвальные сегменты закладываются обычно одновременно на начальной стадии метаморфоза личинок, а лостларвальные сегменты последовательно и постепенно возникают из предпигидиальной эктодермальной зоны роста, причем целом в ларвальных сегментах всегда формируется из мезодермальных полосок путем их разделения на мезодермальные блоки, которые затем становятся полыми.
1 При описании и на рисунках червей сегменты обозначаются римскими цифрами
Число ларвальных сегментов варьирует: у Polynoidae их 8—10, а у Siga-lionidae — 4—6. Большое и изменчивое количество ларвальных сегментов указывает на примитивный тип метаморфоза. Ларвальные сегменты в отличие от постларвальных не образуют половых желез, не имеют целомодуктов и хлора-гогенных клеток. Иногда ларвальные сегменты подвергаются слиянию, образуя перистомиум: например, крупный перистомиум нереид представляет собой продукт слияния трех передних ларвальных сегментов. Вторичная гетерономность относится к постларвальным сегментам, она целиком обусловлена особенностями обитания взрослых червей (адаптивные приспособления).
Количество сегментов у чешуйчатых червей, как и у большинства других полихет, сильно варьирует. Существуют виды, у которых сегментов относительно немного и их число строго постоянное (например, Lepidonotus — всего 26 сегментов), но некоторые виды (многие Lepidasthenia, Arctonoe и др.) состоят из 100 и более сегментов, причем количество последних увеличивается с возрастом и возникновение новых сегментов на заднем конце тела (в предпиги-диальной зоне роста) не прекращается с наступлением половой зрелости. Самые задние сегменты обычно меньше (тоньше и короче) остальных и имеют недоразвитые параподий. Вполне естественно, что черви с большим количеством сегментов одновременно и наиболее длинные. Однако, как уже отмечалось ранее, количество сегментов не всегда строго коррелирует с размерами самих червей. Неопределенно большое количество сегментов у полиноидных червей мы рассматриваем как примитивное состояние, а стабилизацию количества сегментов — как прогрессивный момент. Подсчет сегментов легче всего производить с брюшной стороны, так как спинная сторона червей нередко замаскирована различными придатками, в частности у чешуйчатых червей элитрами. Количество сегментов у представителей афродитид (Aphro-f dita, Laetmonice, Hermonia, Pontogenia) не превышает 50 (колеблется в пределах 32—47). Среди полипоид по числу сегментов выделяется три группы родов: 1) сегментов 25— 30 — Lepidonotus, Hermenia, Euphione, Iphione; 2) сегментов 30—50 — Harmothoe, Lagisca, Eunoe, Antinoe, Gattyana, Bylgides, Eucranta, Malm-grenia, Alentia и др. и 3) сегментов свыше 50 (до 100 и более) — Perolepis, Arctonoe, Lepidasthenia и др. Для большинства сигалионид характерно более 50 сегментов.
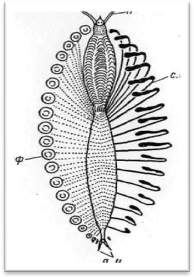
Рис.2. Схема строения пищеварительного канала
Помимо основных наружных перетяжек, отделяющих соседние сегменты друг от друга, в пределах одного сегмента преимущественно на его спинной стороне нередко наблюдается добавочная вторичная, или так называемая ложная кольчатость — псевдосегментация (annuli). У рассматриваемых нами чешуйчатых червей она имеет характер небольших метамерно повторяющихся бороздок или складок, к которым приурочены ресничные пояски и поперечные пигментные полосы, придающие некоторым полиноидам весьма пеструю окраску.
При беглом взгляде на чешуйчатых червей нередко создается впечатление, что они обладают гомономной сегментацией, т. е. на всем протяжении тела у них все сегменты однородны. Такое впечатление объясняется тем, что у многих полиноид вся их спинная сторона как бы равномерно прикрыта спинными чешуйками (элитрами). Однако это ложное представление, так как элитры имеются далеко не на всех сегментах. У чешуйчатых червей наблюдается вполне определенное чередование усиковых сегментов, лишенных элитр, и чешуйчатых сегментов, снабженных элитрами. Таким образом, говорить о полной гомономности сегментов у них не приходится.
Многократное повторение двух или трех смежных сегментов, имеющих разное морфологическое строение, называется метамерией 2-го порядка.
Метамерия 2-го порядка, создающая видимость гомономности, имеет место не только у чешуйчатых червей. Например, у некоторых силлид более длинные спинные усики нередко чередуются через сегмент с более короткими, а иногда (Parasphaerosyllis) изменяется поочередно и форма самих усиков.
Метамерия 2-го порядка — это лишь один из примеров наиболее простой дифференциации сегментов. Она представляет собой начальную стадию образования гетерономности. Наиболее гомономными червями являются филло-доциды, отчасти нереиды. У большинства полихет, в особенности у седентарных, ведущих тубикольный образ жизни, процесс возникновения гетерономности идет значительно дальше и видоизмененные сегменты не чередуются между собой, а приурочены к различным участкам тела, выполняющим разные функции. В связи с этим у некоторых полихет резко обособляются передний участок тела — торакс (так называемые грудные сегменты) и задний участок — абдомен (брюшные сегменты), а иногда и дополнительный хвостовой участок. Наиболее сложная гетерономия наблюдается у ряда седентарных полихет, в частности у Chaetopteridae и Sabellariidae, что обусловлено тубикольным образом жизни. У чешуйчатых червей разделения тела на торакс и абдомен нет, однако XXIII сегмент у них все же играет какую-то роль, отделяя переднюю часть тела со строго закономерным расположением элитр через один сегмент от задней, где расположение элитр еще не стабилизировалось.
У всех чешуйчатых червей, как и у остальных полихет, различают три основных отдела тела. Во-первых, передний, или так называемый головной, отдел, слагающийся из простомиума, или головной лопасти, и двух передних сегментов — перистомиального и буккального. Во-вторых, туловищный отдел (метастомиум), состоящий из большого количества повторяющихся сегментов с развитыми параподиями; этот отдел является наиболее крупной частью червя. В-третьих, анальная лопасть, или пигидий, находящийся на самом заднем конце тела и представляющий собой неметамерный отдел, который образуется у личинок еще до метаморфоза. Впереди пигидия находится зона роста, где возникают новые сегменты. Анальная лопасть обычно снабжена двумя нитевидными анальными усиками, или уритами. У Aphrodita уриты отсутствуют, но на параподиях последних сегментов имеются удлиненные спинные усики, образующие бахрому вокруг ануса.
2.2.1 ГОЛОВНОЙ ОТДЕЛ
Передний, или головной, отдел тела состоит из простомиума (prostomium), иначе головной лопасти, и расположенного за ним первого (перистомиального) и второго (буккального) сегментов (рис. 3, 4). Первый сегмент сильно редуцирован со спины, и от него сохраняются лишь направленные вперед подиаль-ные выросты с перистомиальными, иначе тентакулярными усиками (cirri peri-stomiales, cirri tentaculares). Второй сегмент отличается от последующих туловищных сегментов в основном наличием очень длинных брюшных усиков, которые обозначаются как ротовые (cirri buccales), так как они расположены по бокам ротового отверстия и непосредственно участвуют в обнаружении и захвате пиши. Второй сегмент у всех чешуйчатых червей снабжен первой парой элитр, или спинных чешуек. На переднем крае II сегмента нередко имеется полукруглая складка, или вырост, заходящий сзади на спинную сторону головной лопасти (так называемая затылочная, или нухальная складка). Преобразования I и II сегментов в единый перистомиум путем их слияния у чешуйчатых червей не наблюдается. В этом отношении процессы цефализации у поли-ноид стоят на более низком уровне, чем у нереид, имеющих хорошо выраженный перистомиум в виде широкого ошейника, который представляет собой результат слияния трех передних ларвальных сегментов.
Простомиум у всех чешуйчатых червей хорошо развит и имеет крупные размеры. В процессе метаморфоза личинки он образуется непосредственно из верхнего полушария трохофоры (эписферы), т. е. имеет независимое от всех остальных сегментов происхождение. В соответствии с этим головная лопасть всегда располагается впереди ротового отверстия, находящегося на брюшной стороне червя. У чешуйчатых червей простомиум обычно вытянутой или округлой формы, иногда ширина его может быть больше длины. У переднего края головной лопасти находится 1—3 головных щупальца, или антенны (antennae): одно медиальное и два латеральных.
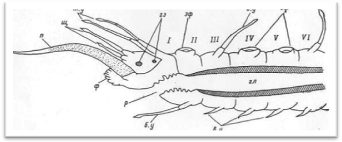
Рис. 3. Продольный разрез через передний отдел тела полиноиды.
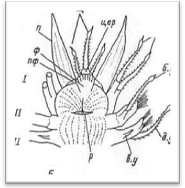
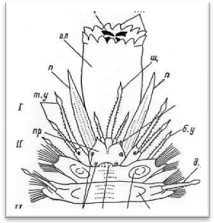
Рис. 4. Головной отдел полиноиды: вид сверху (А) и вид снизу (Б).
Головные щупальца веретенообразной формы, гладкие или с многочисленными мелкими папиллами. Щупальца большей частью имеют обособленные подставки, которые обозначаются как цератофоры, а сами щупальца — цератостили. Наличие цератофор обеспечивает большую подвижность самих щупалец. При медиальном щупальце иногда наблюдаются своеобразные ложковидные органы с ресничным эпителием — так называемые ктенидии (Laetmonice, Sthenelais и др.); назначение этих чувствительных органов точно не выяснено. Количество и характер расположения головных щупалец имеет большое систематическое значение: на этом основании в пределах семейства Polynoidae выделяются несколько подсемейств. У полиноид различают следующие типы головной лопасти (рис.5).
1.Лепидонотоидный тип – три головных щупальца; боковые дольки головной лопасти сильно вытянуты вперед и непосредственно (без базальных подставок) переходят в латеральные щупальца (терминальное положение латеральных щупалец); медиальное щупальце отходит от крупной базальной подставки (цератофора), расположенной в промежутке между боковыми выростами головной лопасти, и находится в одной плоскости с латеральными щупальцами.
2.Арктоноидный тип – три головных щупальца; латеральные щупальца имеют базальные подставки, отходящие от переднего края головной лопасти или несколько сдвинутые на брюшную сторону (субтерминальное положение);
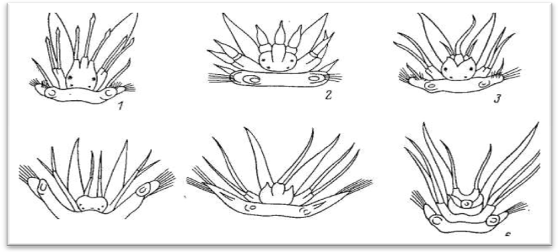
Рис.5 Основные типы головного отдела полиноид. 1 – лепидонотоидный, 2 – арктоноидный, 3 – хармотоидный, 4 – ифионоиднный, 5 – мацеллицефалоидный, 6 - адметеолоидный.
Подставка медиального щупальца лежит поверх латеральных. Арктоноидный тип является промежуточным между лепидонотоидным и хармотоидным.
3.Хармотоидный тип — три головных щупальца; латеральные щупальца с обособленными подставками - имеют вентральное положение, медиальное щупальце с крупной базальной подставкой сдвинуто на дорсальную сторону головной лопасти; дольки головной лопасти нередко с заостренными выростами, так называемыми лобными рогами.
4.Ифионоидный тип — имеются лишь латеральные щупальца, сидящие на боковых выростах головной лопасти; вместо медиального щупальца в задней части головной лопасти под нухальной складкой находится небольшая папилла.
5.Мацеллицефалоидный тип — имеется лишь одно хорошо выраженное медиальное щупальце, сильно сдвинутое на дорсальную сторону головной лопасти и сидящее на крупной базальной подставке (цератофоре); дольки головной лопасти округлые или с небольшими нитевидными придатками, которые представляют собой рудименты латеральных щупалец.
6.Адметеллоидный тип — три головных щупальца; латеральные щупальца лепидонотоидного типа, медиальное щупальце сдвинуто на спинную сторону головной лопасти, при нем имеются особые придатки — так называемые антеннальные чешуйки.
Головная лопасть у всех чешуйчатых червей со спины прикрывает первый (перистомиальный) сегмент, который из-за этого не имеет четких границ. При тентакулярных усиках первого сегмента, иногда обозначаемых как щупальцевидные усики, нередко имеются ацикула и пучок щетинок. Головные щупальца и тентакулярные усики внешне могут мало различаться, однако головные щупальца иннервируются от переднего отдела головного мозга, а перистомиальные усики — от коннективов ротового нервного кольца.
Помимо головных щупалец и тентакулярных усиков, у всех чешуйчатых червей имеются так называемые щупики, или пальпы (palpi), по своей массивности превосходящие все остальные головные придатки. Пальпы отходят с брюшной стороны головного отдела червя, по бокам от ротового отверстия, и внутрь их может заходить целомическая полость. Пальпы имеют очень небольшие подставки — пальпофоры, придающие им дополнительную подвижность. Вдоль наружной поверхности пальп обычно наблюдаются многочисленные продольные ряды мягких чувствительных папилл.
Все указанные здесь головные придатки являются осязательными органами и играют определенную роль при обнаружении пищи, но основным орудием захвата пищи служит выпячивающаяся наружу мускулистая глотка (pharynx), вооруженная нередко мощными хитиноидными челюстями.
На дорсальной стороне головной лопасти расположены глаза; они в большинстве случаев простого типа, без глазных стебельков (омматофоров). Глазные пятна темного (почти черного) цвета, округлой или овальной формы. У полиноид их две пары, причем первая (передняя) несколько крупнее задней, которая к тому же может быть частично прикрыта спинной складкой второго (буккаль-ного) сегмента. Первая пара глаз располагается по бокам головной лопасти в наиболее расширенной ее части или у переднего края, они могут частично переходить и на вентральную сторону головной лопасти (например, у Иаг-mothoe imbricata).

Рис.6. Головной отдел у представителей различных семейств Aphroditiformia.
1— Aphrodita (Aphroditidae), 2 — Harmothoe (Polynoidae), 3 — Stherwlepis (Sigalionidae), 4 — Eulepeihus (Eulepethidae),.7 — Chrysopetalum (Chrysopetalidae), 6 — Pisione (Pisianidae), 7 — Polyodontes (PolyadontidaeJ.
Глазные пятна эпителиального характера и состоят из одного слоя светочувствительных клеток, имеющих сильно вытянутую призматическую форму с темным пигментом в средней своей части. От каждой призматической клетки отходит небольшое нервное волоконце, которое, переплетаясь с соседними, образует оптический нерв, связанный с головным мозгом. Сверху светочувствительные клетки (ретина) прикрыты прозрачной кутикулой (иногда утолщенной), причем у ряда форм под кутикулой наблюдается вдавление или «чаша» заполненная «стекловидным телом» (секретом клеток ретины). Стекловидное тело представляет собой светопреломляющую линзу, или хрусталик (диоптрический аппарат). Иногда под кутикулой дополнительно прослеживается слой эпидермиса и «чаша» преобразуется «в пузырек», который еще более глубоко опускается внутрь мышечных тканей головной лопасти. Наличие светопреломляющих линз хорошо распознается при наружном осмотре, они выглядят в виде светлого кружка в центре темного глаза. Такие линзы наблюдаются преимущественно у пелагических форм, которым требуется обозревать пространство в разных направлениях (например, Lagisca hubrechti). У червей, обитающих на больших глубинах, наоборот, отмечается редукция глаз (Наг-mothoe derjugini, Weberia, Admetella, различные представители мацеллицефалин). У представителей Polyodontidae, живущих в длинных трубках, глаза, как правило, сильно гипертрофированы и находятся на передней овальной стороне вытянутых омматофоров — «глазных ножек». Последние похожи на «рачьи глаза» и служат для быстрой ориентировки, когда червь высовывается из трубки. Наиболее характерные примеры строения головного отдела для различных семейств Aphroditiformia приведены на (рис. 6).
2.2.2 ГЛОТКА И ЕЕ ВООРУЖЕНИЕ
Глотка (pharynx) - мускулистая часть передней кишки, способная выпячиваться наружу через ротовое отверстие. У полихет различают два основных типа глотки: 1) мешковидная глотка (ventral proboscis) в виде относительно короткого слепого мешка (buccal bulb, bulbe pharyngien), расположенного под пищеводом, и 2) трубчатая глотка (axial proboscis, buccal tube), представляющая собой подвижную трубку, постепенно переходящую в среднюю кишку. Первый, весьма специализированный тип глотки среди эррантных полихет характерен лишь для эвницид и амфяномид; наиболее сложно устроена мешковидная глотка эвницид, вооруженная нижними и верхними челюстями (мандибулой и максиллами). У чешуйчатых червей глотка трубчатая. Механизм выпячивания сильно мускулистой глотки чешуйчатых червей такой же, как у других полихет с этим типом глотки, т. е. глотка выпячивается наружу наподобие пальца перчатки. Выпячивание происходит вследствие давления полостной жидкости и регулируется специализированными мышцами. Выпяченная глотка у некоторых видов достигает трети длины самого червя.
У наружного отверстия выпяченной глотки полиноид обычно имеются отдельные крупные округлые, мягкие краевые (оральные) папиллы или многочисленные удлиненные сосочки, образующие густую наружную бахрому.
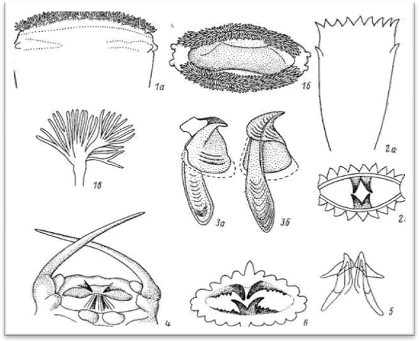
Рис. 7. Глотка и челюстной аппарат.
1—передний край выпяченной глотки Laetmonice: вид сбоку (а), вид спереди (б) и отдельная краевая па-шглла (в); 2 — глотка Leyidonotus; вид сбоку (а) и вид спереди (б); 3 — отпрепарированные челюстные крючки Lepidasthenm: дорсальный правый (а) и вентральный левый (б); 4 — передняя часть глотки Macellicephaloides; 5 — челюстной аппарат Рisione; 6 — челюстное вооружение Polyodontes, вид глотки спереди (3, б).
Краевые папиллы богаты железами, выделяющими ферменты, которые участвуют в пищеварении. Наружная стенка выпяченной глотки обычно гладкая, без папилл. Примеры строения глотки и ее вооружения у чешуйчатых червей приведены на (рис.7).
Для большинства полиноид характерно наличие как на дорсальной, так и на вентральной стороне глотки по 9 крупных краевых папилл. Иногда их может быть несколько больше, а именно 11+11 (Lepidasthenia) и даже до 14—22 пар (Hololepida). У рода Hermilepidonotus описан крайне необычный для полиноид характер краевых папилл: их до 30 пар, причем они не простой треугольной формы, а имеют с внутренней стороны до 20 добавочных пальцевидных отростков, образующих у входного отверстия как бы щетку, напоминающую таковую Aphroditidae s. str. (Laetmonice) (рис. 7, 1). Несколько необычен характер папилл у мацеллицефалин: так, представители рода Macellicephaloides (рис. 7, 4) имеют на глотке две очень длинные боковые усиковидные папиллы, которые в морфологическом и функциональном отношениях весьма сходны с боковыми усиками на глотке пелагических альциопид.
Челюстной аппарат, представляющий собой локальные утолщения гиподермы, у чешуйчатых червей развит различно. Наиболее мощные челюсти в виде четырех крупных крючков (или когтей) наблюдаются у большинства полиноид, сигалионид и полиодонтид. При выпяченной глотке эти крючки частично выставлены наружу, располагаясь попарно на вентральной и дорсальной сторонах глотки. Каждый челюстной крючок имеет несколько расширенное основание и слегка оттопыренную утонченную боковую пластинку, или крылышко, так называемый элерон (aileron). Среди аберрантной группы преимущественно глубоководных полиноид — Macellicephalinae — наблюдается постепенная редукция челюстного аппарата: Macellicephala, так же как и все полиноиды, имеют 2 дорсальных и 2 вентральных крючка; Macellicephaloides — 1 дорсальный и 2 вентральных крючка; у Macelloides имеются лишь очень тонкие, слегка зазубренные утолщения по наружному краю глотки, a Macella вообще лишена на глотке каких-либо следов хитиноидных утолщений. Незначительно развит челюстной аппарат у представителей Eulepethidae. Для Aphroditidae s. str. характерно отсутствие челюстных образований. Глотка Chrysopetalidae снабжена лишь двумя небольшими стилетами.
Челюсти чешуйчатых червей, как и других полихет, состоят в основном из; кератина, или рогового вещества; они обычно темного, почти черного цвета. На некоторых челюстных пластинках хорошо видны так называемые годовые кольца, по которым можно судить о возрасте червей. Детали строения челюстей имеют значение для таксономии — диагностики родов и семейств, а также для филогенетических построений. Челюсти хорошо сохраняются в ископаемом состоянии. В онтогенезе челюсти у полиноид закладываются уже на второй стадии нектохеты.
Челюстной аппарат полихет выполняет различные функции. В довольно редких случаях он используется как орудие защиты и нападения (последнее было отмечено среди нереид). Глотка может играть существенную роль при ло-комоции и зарывании в грунт, в частности при постройке ходов — ranepefr в толще грунта; в этих случаях она действует как вспомогательное «якорное»-устройство (например, у глицерид и нефтиид).
Наличие или отсутствие челюстного аппарата у полихет еще ничего не говорит о том, являются ли данные черви плотоядными или детритоядными животными. Известно, что многие полихеты, лишенные челюстного аппарата, ведут хищнический образ жизни, например филлодоциды, и, наоборот, некоторые многощетинковые черви с развитыми челюстями используют последние в основном как грабли для сбора детрита, в частности некоторые нереиды. В желудках большинства полиноид, снабженных мощными челюстями, обычно обнаруживают не только мелких животных, но и различного рода растительные обрывки и разнообразный детрит. По всей видимости, полиноиды, как и большинство других полихет, всеядные животные. Челюстной аппарат полихет, как правило, не служит жевательным органом, а лишь служит для схватывания и удерживания добытой пищи, которая затем путем втягивания глотки поступает в кишечник. У основания челюстей глицерид и некоторых нереид имеются особые ядовитые железы, их яд умертвляет жертву. У полиноид такие железы не описаны, но наличие их вполне возможно.
2.2.3 ПАРАПОДИАЛЬНЫЙ КОМПЛЕКС
Все сегменты метастомиума, иначе туловищные сегменты, снабжены боковыми выростами — параподиями, вооруженными многочисленными щетинками. Пара-подии в основном служат локомоторными органами, но одновременно в не-которых случаях выполняют и дыхательные функции, а также являются орудиями защиты. Более подробно остановимся на морфологии параподий Aphroditiformia, у которых, как и у большинства эррантных полихет, параподий моностикного характера, т. е. невроподии и нотоподии объединены общим подиальным стволом (в противоположность многим седентарным полихетам, у которых имеются дистикные параподий, характеризующиеся разобщенными ветвями).
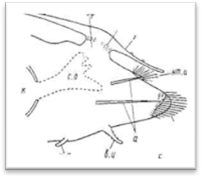
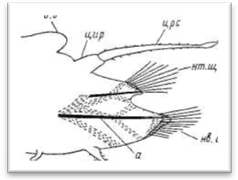
Рис. 8. Усиковая (А) и чешуйчатая (Б) параподии полиноид.
Параподий чешуйчатых червей имеют большей частью сильно вытянутую форму и четко отделены от главного ствола тела самого червя (на брюшной стороне они ограничиваются нефридиальными папиллами). Длина параподий нередко равна ширине тела и даже может ее превосходить. Различают два основных типа параподий: 1) усиковые параподии, снабженные удлиненным верете-образной формы спинным усиком, выполняющим осязательную функцию, и 2) чешуйчатые параподии, у которых спинной усик заменен плоской чешуйкой, или элитрой, выполняющей преимущественно защитную функцию (рис. 8). Параподии снабжены двумя крупными внутренними осевыми щетинками, как правило, темного цвета. Это так называемые ацикулы (aciculum), которые соответствуют двум ветвям параподий: верхней, или спинной (нотоподия— notopodium), и нижней, или брюшной (невроподия — neuropidium). Такие параподии обозначаются как двуветвистые (biramus, biremis), однако верхняя ветвь нередко остается недоразвитой и выражена лишь небольшим бугорком на передне-верхней стороне подиального ствола. Параподии с недоразвитой нотоподией иногда называются «ложнодвуветвистые» или «полуразветвленные» (sub-biramus, sesquiramus). При беглом взгляде последние могут быть приняты за одноветвистые; они наблюдаются преимущественно лишь у многосегментных полиноид типа Arctonoe, Lepidasihenia и Perolepis. Короткие полиноиды — Harmotkoe и большинство других родов имеют, напротив, мощную нотоподию, снабженную большим колич