


Меристемы или образовательные ткани
МЕРИСТЕМЫИЛИ ОБРАЗОВАТЕ ЛЬНЫЕ ТКАНИ
Цитоплазма. Химический состав, физическое состояние, строение и свойства. Поступление веществ в клетку. Типы движения цитоплазмы.
Цитоплазма бесцветная, имеет слизистую консистенцию и содержит различные вещества, в том числе и высокомолекулярные соединения, например белки, присутствие которых обусловливает коллоидные свойства цитоплазмы. Цитоплазма – часть протопласта, заключенная между плазмалеммой и ядром. Основу цитоплазмы составляет ее матрикс, или гиалоплазма, - сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель.
В цитоплазме растительных клеток имеются органоиды: небольшие тельца, выполняющие специальные функции, - пластиды, комплекс Гольджи, эндоплазматическая сеть, митохондрии и т.д. В цитоплазме осуществляется большая часть процессов клеточного метаболизма, исключая синтез нуклеиновых кислот, происходящих в ядре. Цитоплазма пронизана мембранами – тончайшими (4-10нм) пленками, построенными в основном из фосфолипидов и липопротеинов. Мембраны ограничивают цитоплазму от клеточной оболочки и вакуоли и внутри цитоплазмы образуют эндоплазматическую сеть (ретикулум) – систему мелких вакуолей и канальцев, соединенных друг с другом.
Важнейшим свойством цитоплазмы, связанным прежде всего с физико-химическими особенностями гиалоплазмы, является ее способность к движению. В клетках с одной крупной вакуолью движение осуществляется обычно в одном направлении (циклоз) за счет особых органоидов – микрофиламентов, представляющих собой нити особого белка - актина. Движущая гиалоплазма увлекает пластиды и митохондрии. Клеточный сок, находящийся в вакуолях, представляет собой водный раствор различных веществ: белков, углеводов, пигментов, органических кислот, солей, алкалоидов и т.п. Концентрация веществ, находящихся в клеточном соке, обычно выше концентрации веществ во внешней среде (почва, водоемы). Различие концентраций в известной мере обусловливает возможность поступления в клетку воды и почвенных растворов, что до некоторой степени объясняется явлением осмоса. В клетке роль полупроницаемой мембраны играет цитоплазма. Пограничные слои цитоплазмы, выстилающие оболочку и клеточную вакуоль, проницаемы только для воды и некоторых растворов, но непроницаемы для многих растворенных в воде веществ. Это свойство цитоплазмы получило название полупроницаемости или избирательной проницаемости. В отличие от цитоплазмы клеточная оболочка проницаема для всех растворов, непроницаема она только для твердых частиц. Поступление веществ в клетку нельзя сводить только к осмотическим явлениям, которые выражены во взрослых клетках с хорошо развитыми вакуолями. В действительности это очень сложный процесс, обусловленный многими факторами. Активное участие в поглощении веществ принимает вся система коллоидов цитоплазмы. Интенсивность движения зависит от температуры, степени освещения, снабжения кислородом и т.д.
В очень молодых клетках цитоплазма заполняет почти всю их полость. По мере роста клетки в цитоплазме появляются мелкие вакуоли, заполненные клеточным соком, представляющим собой водный раствор различных органических веществ. Впоследствии, при дальнейшем росте клетки, вакуоли увеличиваются в размерах и, сливаясь, часто образуют одну большую центральную вакуоль, оттесняющую цитоплазму к оболочке клетки. В таких клетках все органоиды располагаются в тонком постенном слое цитоплазмы. Иногда ядро остается в центре клетки. В этом случае цитоплазма, образующая вокруг него ядерный кармашек, соединяется с постенным слоем тонкими цитоплазматическими тяжами.
В слое цитоплазмы расположены хлоропласты, выстилающие верхнюю стенку. Они представляют собой почти округлые или слегка овальные тельца. Изредка можно встетить пластиды, перетянутые посередине.
Понятие о тканях. Классификация тканей. Отличие постоянных тканей от образовательных.
У большинства наземных растений клетки тела неодинаковы по своим функциям, строению и происхождению. Эта разнородность возникла и закрепилась, в процессе приспособления растений к различиям воздушной и почвенной сред. Системы клеток, структурно и функционально сходны друг с другом и обычно имеющие общее происхождение, получили название ткачей. Ткани имеются почти у всех высших растений. Нет их только у части моховидных (печеночники). Водоросли и багрянки (низшие растения), как правило, также не имеют развитых тканей.
Комплексы клеток, сходных по функциям, а большей частью и по строению, имеющих одинаковое происхождение и определенную локализацию в теле растения, называют тканями.
Распределение тканей в органах растений и их структура тесно связаны с выполнением ими определенных физиологических функций.
Ткани, состоящие из одного типа клеток, получили название простых, а состоящие из разных типов клеток — сложных, или комплексных. Существуют различные классификации тканей, но все они достаточно условны. Растительные ткани делят на несколько групп в зависимости от основной функции:
1) меристемы, или образовательные ткани (ткани состоящие из живых тонкостенных, интенсивно делящихся клеток);
а) верхушечные (апикальные) меристимы (расположенны на верхушках стеблей и в окончаниях корней) обусловливают рост этих органов в длину;
б) боковые мерестимы – камбий и феллоген (камбий обеспечивает утолщение стебля и корня. Феллоген образует пробку)
2) покровные (защищают внутренние ткани растений от прямого влияния внешней среды, регулируют испарение и газообмен)
а) эпидермис;
б) пробка;
3) проводящие (обеспечивают проведение воды, почвенных растворов и продуктов ассимиляции, вырабатываемых листьями. Проводящие ткани по происхождению могут быть первичными и вторичными.);
а) кселима или древесная ткань (ткань проводящая воду)
б) флоэма или луб (ткань проводящая органические вещества, образованные растением в процессе фотосинтеза);
4) механические (обусловливают прочность растения);
а) коленхима (состоит из паренхимы или несколько удлиненных клеток с неравномерно утолщенными целлюлозными стенками);
б) склеренхима (клетки имеют равномерно утолщенные одревесневшие стенки);
1) волокна;
2) склериды;
5) основные (состоящие из однородных паренхимных клеток, которые заполняют пространство между другими тканями);
6) секреторные, или выделительные (содержащие продукты отброса).
Лишь клетки меристематических тканей способны к делению. Клетки прочих тканей, как правило, к делению неспособны, и их число увеличивается за счет деятельности соответствующих меристем. Такие ткани называют постоянными. Постоянные ткани возникают из меристем в результате клеточной дифференцировки. Дифференцировка заключается в том, что в ходе индивидуального развития организма (онтогенеза) возникают качественные различия между первоначально однородными клетками, при этом изменяются строение и функциональные свойства клеток. Обычно дифференцировка необратима. На ход ее оказывают влияние вещества, выполняющие роль гормонов.
МЕРИСТЕМЫИЛИ ОБРАЗОВАТЕ ЛЬНЫЕ ТКАНИ
Меристемы (от греч. «меристос» — делимый), или образовательные ткани, обладают способностью к делению и образованию новых клеток. За счет меристем формируются все прочие ткани и осуществляется длительный (в течение всей жизни) рост растения. У животных меристемы отсутствуют, чем объясняется ограниченный период их роста. Клетки меристем отличаются высокой метаболической активностью. Одни клетки меристем, получившие название инициальных, задерживаются на эмбриональной стадии развития в течение всей жизни растения, другие постепенно дифференцируются и превращаются в клетки различных постоянных тканей. Инициальная клетка меристемы принципиально может дать начало любой клетке организма. Тело наземных растений — производное относительно немногих инициальных клеток.
Первичные меристемы обладают меристематической активностью, т. е. способны к делению изначально. В ряде случаев способность к активному делению может вновь возникнуть и у клеток, уже почти утративших это свойство. Такие «вновь» возникшие меристемы называют вторичными.
В теле растения меристемы занимают различное положение, что позволяет их классифицировать. По положению в растении выделяют верхушечные, или апикальные (от лат. «апекс» — верхушка), боковые, или латеральные (от лат. «латус» — бок), и интеркалярные меристемы.
Апикальные меристемы располагаются на верхушках осевых органов растения и обеспечивают рост тела в длину, а латеральные — преимущественно рост в толщину. Каждый побег и корень, а также зародышевый корешок, почечка зародыша имеют апикальную меристему. Апикальные меристемы первичны и образуют конусы нарастания корня и побега (рис. 1).
Латеральные меристемы располагаются параллельно боковым поверхностям осевых органов, образуя своего рода цилиндры, на поперечных срезах имеющие вид колец. Часть из них относится к первичным. Первичными меристемами являются прокамбий и перицикл, вторичными — камбий и феллоген.
Интеркалярные, или вставочные, меристемы чаще первичны и сохраняются в виде отдельных участков в зонах активного роста (например, у оснований междоузлии, в основаниях черешков листьев).
Существуют также раневые меристемы. Они образуются в местах повреждения тканей и органов и дают начало каллюсу — особой ткани, состоящей из однородных паренхимных клеток, прикрывающие место поражения Каллюсо-образовательная способность растений используется в практике садоводства при размножении их черенками и прививками. Чем интенсивнее каллюсообразование, тем больше гарантия срастания подвоя с привоем и укоренения черенков. Образование каллюса— необходимое условие культуры тканей растения на искусственных средах.
Клетки апикальных меристем более или менее изодиаметричны по размерам и многогранны по форме. Межклетников между ними нет, оболочки тонкие, содержат мало целлюлозы. Полость клетки заполнена густой цитоплазмой с относительно крупным ядром, занимающим центральное положение. Вакуоли многочисленные, мелкие, но под световым микроскопом обычно не заметны. Эргастические вещества, как правило, отсутствуют. Пластид и митохондрий мало и они мелки.
Клетки боковых меристем различны по величине и форме. Они примерно соответствуют клеткам тех постоянных тканей, которые из них в дальнейшем возникают. Так, в камбии встречаются как паренхимные, так и прозенхимные инициали. Из паренхимных инициалей образуется паренхима проводящих тканей, а из прозенхимных — проводящие элементы.
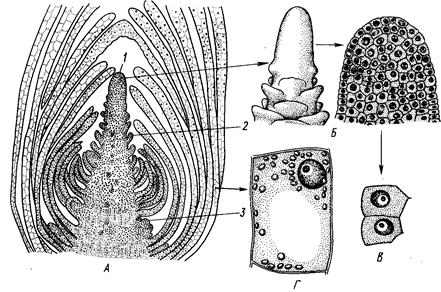
Рис. 1. Верхушечная меристема побега элодеи. А — продольный срез; 5 — конус нарастания (внешний вид и продольный срез); В — клетки первичной меристемы;
Г — паренхимная клетка сформировавшегося листа:
1 ~ конус нарастания, 2 — зачаток листа, 3 — бугорок пазушной почки
Зоны корня. Особенности морфологического строения каждой зоны в связи с выполняемыми функциями. Рост корня в длину и втолщину.
Наряду с побегом корень - основной орган высшего растения, в типичном случае выполняющий функцию минерального и водного питания. Другая важная функция корня, тесно связана с основной, - закрепление, растения в почве.
Различные части корня выполняют неодинаковые функции и характеризуются определенными морфологическими особенностями. Эти части получили название зон (рис. 2). Кончик корня снаружи всегда прикрыт корневым чехликом, защищающим апикальную меристему. Клетки корневого чехлика продуцируют слизь, покрывающую поверхность корня. Благодаря слизи снижается трение о почву, и ее частицы легко прилипают к корневым окончаниям и корневым волоскам.
Корневой чехлик состоит из живых паренхимных клеток, возникающих у большинства однодольных из особой меристемы, получившей название калиптрогена, а у двудольных и голосеменных — из верхушечной меристемы кончика корня. Водные растения корневого чехлика обычно не имеют.
Под чехликом располагается зона деления, представленная меристематической верхушкой корня, его апексом. В результате активности апикальной меристемы формируются все прочие зоны и ткани корня.
Делящиеся клетки сосредоточены в зоне деления, имеющей размеры около 1 мм. Эта часть корня заметно отличается от прочих зон своей желтоватой окраской. Вслед за зоной деления располагается зона растяжения (роста). Она также невелика по протяженности (несколько миллиметров), выделяется светлой окраской и как бы прозрачна. Клетки зоны роста практически не делятся, но способны растягиваться в продольном направлении, проталкивая корневое окончание в глубь почвы. Они характеризуются высоким тургором, что способствует активному раздвиганию частиц почвы. В пределах зоны роста происходит дифференциация первичных проводящих тканей.
Окончание зоны роста заметно по появлению на эпиблеме многочисленных корневых волосков. Корневые волоски располагаются в зоне—всасывания, функция которой понятна из ее названия. На корне она занимает участок от нескольких миллиметров до нескольких сантиметров. В отличие от зоны роста участки этой зоны уже не смещаются относительно частиц почвы. Основную массу воды и растворов солей молодые корни усваивают в зоне всасывания с помощью корневых волосков.
Корневые волоски появляются в виде небольших сосочков — выростов клеток эпиблемы. Рост волоска осуществляется у его верхушки. Оболочка корневого волоска растягивается быстро. По прошествии определенного времени корневой волосок отмирает. Продолжительность его жизни не превышает 10—20 дней.
Выше зоны всасывания, там, где исчезают корневые волоски, начинается зона проведения. Строение этой зоны на разных ее участках неодинаково. По этой части корня вода и растворы солей, поглощенные корневыми волосками, транспортируются в вышележащие отделы растения. Участки зоны проведения занимают фиксированное положение относительно участков почвы, не смещаясь относительно их. Несмотря на фиксированное положение в пространстве конкретных участков зон поглощения и проведения, эти зоны сдвигаются вследствие верхушечного роста. В результате этого всасывающий аппарат постоянно перемещаетсяв почве.
В пределах одной корневой системы существуют корни, выполняющие разные функции. У большинства растений различаются ростовые и сосущие окончания. Ростовые окончания долговечнее, относительно мощные, быстро удлиняются и продвигаются в глубь почвы. Сосущие окончания недолговечны, возникают в большом числе на вростовых корнях
и удлиняются медленно. У деревьев и кустарников выделяют видоизмененные скелетные и полускелетные корни, на которых возникают недолговечные корневые мочки, несущие множество сосущих окончаний.