Разноспоровость у высших растений
МИНИСТЕРСТВО ОБРАЗОВАНИЯ РФ
ЧЕРЕПОВЕЦКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
ИНСТИТУТ ВАЛЕОЛОГИИ
Кафедра биологии и общей экологии
специальность 032400
«РАЗНОСПОРОВОСТЬ У ВЫСШИХ РАСТЕНИЙ»
Курсовая работа
студентки 6Б-21
Тумановой Н.С.
Научный руководитель
Румянцева А.В.
ЧЕРЕПОВЕЦ
2001
СОДЕРЖАНИЕ стр
- Введение……………………………………………………….…2
1.1. Жизненный цикл высших растений…………………………4
- Основная часть……………………………………………..…….3
2.1. Отдел Риниофиты …………………………….…………..…7
2.2. Отдел Зостерофиллофиты… …………………………….….7
2.3. Отдел Моховидные ………………………………..…………7
2.4. Отдел Плауновидные ………………………………………...7
2.5. Отдел Псилотовидные ….…………………………………...10
2.6. Отдел Хвощевидные ………………………………….……..10
2.7. Отдел Папоротниковидные ………………………………...10
2.8. Отдел Голосеменные…………………………………………12
2.9. Отдел Цветковые……………………………………………..14
- Вывод………………………………………………………………18
- Список используемой литературы……………………………….19
Приложение……………………………………..…………20
1. Введение
Тема моей курсовой работы называется «Разноспоровость высших растений». В своей работе я рассматриваю все высшие растения, которые являются разноспоровыми. Высшие растения делятся на девять отделов:
- Отдел Риниофиты (Rhyniophyta)
- Отдел Зостерофиллофиты(Zosterophyllophyta)
- Отдел Моховидные (Bryophyta)
- Отдел Плауновидные (Lycopodiophyta)
- Отдел Псилотовидные (Psilotophyta)
- Отдел Хвощевидные (Equisetophyta)
- Отдел Папоротниковидные (Polypodiophyta)
- Отдел Голосеменные (Pinophyta или Gymnospermae)
- Отдел Цветковые (Magnoliophyta)
Из этих отделов к разноспоровым высшим растениям относят:
- Отдел Плауновидные (Lycopodiophyta)
- Отдел Хвощевидных (Equisetophyta)
- Отдел Папоротниковидные (Polypodiophyta)
- Отдел Голосеменные (Pinophyta или Gymnospermae)
- Отдел Цветковые (Magnoliophyta)
Объектом исследования в этой курсовой работе является разноспоровость как явление, заключающееся в существовании мегаспор и микроспор. Микроспорой называется мелкая спора высших разноспоровых растений, из которой развивается мужской гаметофит (половое поколение); у высших споровых растений из микроспоры развивается мужской заросток, у семенных растений микроспорами являются пыльцевые зерна (пылинки). Мегаспора (макросора), более крупная из спор, образуемых высшими разноспоровыми растениями, из которой развивается женский гаметофит(половое поколение); у высших споровых растений мегаспора, прорастая, дает начало женскому заростку (у семенных растений он называется зародышевым мешком). Микроспоры, более мелкие, образуются в микроспорангии, а мегаспоры, более крупные, развиваются в мегаспорангиях. Каждая микроспора дает начало одному мужскому гаметофиту, а из каждой мегаспоры образуется по одному женскому гаметофиту. Поэтому некоторые авторы, например Б. М. Козо-Полянский, называют микроспоры «мужскими спорами», а мегаспоры – «женскими спорами».
В своей курсовой работе я ставлю цель выделить отделы высших растений, относящихся к разноспоровым. Также выделить особенности образования мегаспор и микроспор, их размеры и строение.
Для написания этой курсовой работы я пользуюсь основным литературным источником – «Жизнь растений» (4 том, 5 том 1часть). В этой книге достаточно полно освещены вопросы, необходимые для написания курсовой работы. Более полно явление разноспоровости рассмотрено в отделах Голосеменные и Цветковые. В общей характеристики этих отделов четко обозначены различия микроспор и мегаспор в их строении, местоположении и развитии. Таким образом основными задачами курсовой работы является обоснование всех факторов разноспоровости высших растений, выявлении особенности разноспоровости. Для решения задач поставленных в моей курсовой работе по изучению явления разноспоровости высших растений необходимо знать их жизненный цикл.
1.1. Жизненный цикл высших растений
Жизненный цикл высшего растения состоит из двух фаз (или поколений) – бесполой, или спорофазы (спорофита), и половой, или гаметофазы (гаметофита). Началом развития полового поколения является спора, которая, как нам уже известно, образуется внутри спорангии. В спорангии может образовываться самое различное число спор. У равноспоровых высших растений, у которых споры еще не дифференцированы на микроспоры и мегаспоры, в каждом спорангии обычно образуется не менее 32 спор и лишь очень редко только 8 спор.
У разноспоровых высших растений число микроспор в каждом микроспорангии также обычно не бывает меньше 32, но в мегаспорангии, как правило, образуется только одна мегаспора.
Развитие спор внутри спорангии, особенно развитие мегаспор, происходит довольно сложным путем. Внутри спорангии закладывается первичная спорообразующая ткань. В результате нескольких митотических делений всех спорогенные клетки или только часть их дают начало материнским клеткам спор, или спороцитам. Из каждого спороцита в результате мейоза может образовываться 4 споры. Число хромосом в спорах по сравнению со спороцитом в двое меньше, что объясняется редукцией числа хромосом, происходящей в процессе мейоза следовательно, именно здесь происходит смена ядерных фаз, переход спорофазы в гамето фазу.
Микроспоры и мегаспоры окружены оболочкой (спородермой). Спородерма выполняет разнообразные функции, связанные с жизнедеятельностью споры. На ранних стадиях развития оболочка обеспечивает избирательное поступление веществ из полости спорангия (или гнезда пыльника) в развивающуюся спору. Покидая материнское растение, споры некоторое время существуют самостоятельно. В этих условиях на их оболочку, не прикрытую другими клетками, ложится полная ответственность за сохранение содержимого от высыхания, чрезмерного увлажнения или механических повреждений. Кроме этого, являясь наружным скелетом, спородерма сохраняет определенную форму.
Большое количество жизненно важных функций, ложащихся на оболочку всего лишь одной клетки, вызвало необходимость постоянного совершенствовать строение спородермы. В результате выработалась чрезвычайно прочная, устойчивая к высоким температурам, химическим реагентам и воздействию микроорганизмов, совершенная по конструкции, форме и скульптуре оболочка. Особенностью наружных слоев спородермы является также их поистине удивительная сохранность в ископаемом состоянии.
Существенным признаком в строении спородермы являются также ее форма, определяющаяся многими факторами: соединением спор в тетраде, расположением апертур, свойствами каллозы и т. д. Для бессеменных растений наиболее характерны споры, имеющие тетраэдрическую (плауны, многие папоротники, мхи) или бобовидную, переходящую в эллипсовидную (многие папоротники) форму. Хвощи и большинство моховидных имеют шарообразные споры. Голосеменные унаследовали эллипсовидную и шарообразную формы, в некоторых линиях эволюции трансформировав ее путем образования воздушных мешков.
Спородерма современных растений составлена из нескольких хорошо различимых слоев, имеющих различное происхождение и строение. Самый наружный слой – перина – характерен для многих спор мхов, папоротников, хвощей. В виде фрагментов образуется у некоторых голосеменных и, как исключение, представляет самый верхний слой спородермы некоторых очень примитивных цветковых растений. Он формируется из структур, образующихся в клетках стенках спорангия или гнезда пыльника. Следующий слои спородерма – экзина и интина, образуются протопластом самой споры.
Морфологические признаки слоев спородермы формируются постепенно в процессе развития спор. Сразу же после мейоза вокруг каждой споры образуются временная оболочка – каллоза.
Содержимое споры представляет собой первую клетку гаметофазы, и из нее в результате ряда митотических делений развивается гаметофит. Каждый изоспора дает начало обоеполому многоклеточному гаметофиту, на котором развиваются многоклеточные половые органы – антеридии и архегонии. Из гетероспор также развиваются гаметофиты, но содержимое микроспор дает начало только мужским гаметофитам, а содержимое мегаспор – женским гаметофитам.
Рассмотрев, жизненный цикл высших растений я отметила особенности как равноспоровых, так и разноспоровых растений.
2. Основная часть
2.1. Отдел Риниофиты (Rhyniophyta)
Риниофиты (Rhyniophyta) относят к равноспоровым растениям. В спорангиях риниофитов развивались многочисленные споры, по строению оболочки типичные для высших растений. Споры развивались в тетрадах и были снабжены трехлучевым тетрадным рубцом. Раскрывание оболочки такой споры происходило вдоль лучей тетрадного рубца.
2.2. Отдел Зостерофиллофиты (Zosterophyllophyta)
Зостерофиллофиты (Zosterophyllophyta) также относят к равноспоровым растениям. Их спорангии снабжены короткой ножкой и содержат одинаковые споры.
2.3. Отдел Моховидные (Bryophyta)
Моховидные (Bryophyta) тоже являются равноспоровыми растениями. Споры – обычно одноклеточные образования, служащие для бесполого размножения мхов и их распространения. Размеры спор сильно колеблются. Форма спор варьирует незначительно, обычно они шаровидные, реже овальные, иногда слабо округло-угловатые, редко почковидные.
2.4. Отдел Плауновидные (Lycopodiophyta)
А вот среди плауновидных (Lycopodiophyta) есть как равно-, так и разноспоровые растения. Разноспоровые растения обладают листьями с язычками. Споры плауновидных обычно с трехлучевым рубцом. Гаметофиты разноспоровых плауновидных однополые, незеленые, развиваются обычно в течение нескольких недель за счет питательных веществ, содержащихся в споре, и по достижении зрелости не выступают наружу, за пределы оболочки споры.
Класс изоэтопситы (Isoetopsida) включает разноспоровые плауновидные. Сюда относят порядки: селагинелловые (Selaginellales), лепидодендровые (Lepidodendrales) и полушниковые (Isoetales), причем лепидондровые полностью вымерли, а два других порядка представлены и ныне живущими растениями.
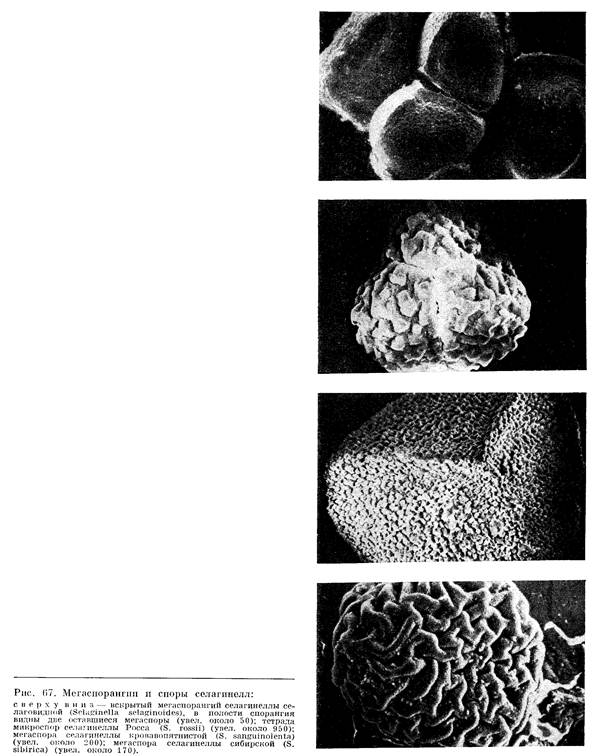
К порядку селягинелловые (Selaginellales) относят разноспоровые многолетние травянистые ветвистые растения с цельными листьями, обладающими язычком, и с придаточными корнями, отходящими обычно от тонкого стебля; некоторые ископаемые представители обладали хорошо развитым ризофором. Спорангии располагаются на стебле близ пазухи листа или в пазухе листа. Спорофиллы собраны в стробилы. Гаметофиты однополые, мелкие, развиваются в течении года, не покидая споровой оболочки, за счет запасов питательных веществ, находящихся в споре. Сперматозоиды двужгутиковые. Современные представители входят в род селягинелла (Selaginella), называемый также по-русски плауном. Основным способом размножения селягинелл является бесполое размножение с помощью спор. Стробилы располагаются на концах боковых ветвей. Они либо довольно резко отграничены от остальной части облиственного побега, либо внешне почти неотличимы от нее. Спорофиллы сидят на оси стробила по спирали или накрест супротивно в 4 продольных рядах.
У большинства видов как мегаспорофиллы с мегаспорангиями, так и микроспорофиллы с микроспорангиями находятся в одном и том же стробиле, при этом первые обычно занимают нижнюю, а вторые – верхнюю его часть, реже они располапфются вдоль оси стробила двумя вертикальными рядами или, наконец, размещены без определенного порядка. Мегаспорангии обычно много крупнее микроспорангиев. Мега- и микроспоры, как правило, с трехлучевым тетрадным рубцом, с многослойной и сложно устроенной оболочкой, скульптура поверхности которой очень разнообразна и является хорошим диагностическим признаком. Диаметр микроспор в зависимости от вида варьирует от 20 до 100 мкм, мегаспор – от 100до 1,5 мм. Споры бывают беловатыми, желтоватыми, красноватыми, буроватыми и почти черными. Раскрывание спорангии происходит иногда столь резко, что микроспоры или тетрады микроспор выбрасываются на расстояние до 2 см, а мегаспоры – на расстояние до 10 см. далее споры рассеиваются ветром или водой.
Например, у разных видов рода полушник (Isetes) в мегаспорангиях развивается от 50 до 2350 мегаспор диаметром от 90 до 900 мкм в микроспорангиях – от нескольких тысяч до миллиона микроспор диаметром от 20 до 40 мкм. В отличие от других споровых растений полушник обладает мега- и микроспорами разного морфологического типа мегасапоры трехлучевые, микроспоры билатеральные, однолучевые. Поверхность как мегаспор, так и микроспор бывает очень разнообразной. Самый внешний слой споровой оболочки мегаспор состоит из кремзема, и по высыхании его споры обычно кажутся белыми. Спорангии полушника не имеют какого-либо механизма для вскрывания, стенка их при ежегодном отмирании листьев сгнивает, и споры высвобождаются. Распространяются споры главным образом водой, но в ряде случаев их переносят дождевые черви, через кишечный тракт которых споры, покрытые панцирем из кремнезема, проходят без повреждений. Не исключена также возможность переноса спор земноводных и наземных видов полушника птицами и копытными животными.
Споры прорастают обычно на следующий год после созревания. Мужские и женские гаметофиты полушника редуцированы в еще большей степени по сравнению с соответевующеми гаметофитами селягинелл. Мужской гаметофит состоит из одной проталлиальной клеткии антеридия с однойслойной стенкой из четырех клеток и с четырмя сперматидиями. При созревании гаметофита оболочка споры лопается и освобожддаются 4 многожгутиковых сперматозоида. При развитии женского гаметофита, как и у селагинелл, отмечается стадия свободных ядерных делений. Спустя некоторое время лопнувшая оболочка споры обнажает поверхность незеленого женского гаметофита с архегониями.
После оплодотворения из зиготы развивается зародыш, состоящий из листа, корня и ножки, с помощью которого зародыш высасывает питательные вещества из крахмалоносной ткани гаметофита.первый лист и первый корень пробивают ткань гаметофита и выходят наружу. Молодой спорофит долго остается прикрепленным к женскому гаметофиту.
2.5. Отдел Псилотовидные (Psilotophita)
Среди ныне живущих высших растений совершенно обособленное положение занимает маленький отдел псилотовидные (Psilotophita), в который входят только равноспоровые растения. Споры псилотовых обячно билатерально симметричные, почковидные, с прозрачными оболочками и имеют сетчатую поверхность.
2.6. Отдел Хвощевидные (Equisetophyta)
Подавляющее большинство хвощевидных (Equisetophyta) – равноспоровые растения, и лишь некоторые ископаемые формы были разноспоровые.
2.7. Отдел Папоротниковидные (Polypodiophyta)
В отделе папоротниковидные (Polypodiophyta) только один подкласс сальвинииды (Salviniidae) включает в себя разноспоровые растения. Десять видов современных сальвиний (Salvinia) являются остатками некогда крупного рода, возникшего по-видимому, в мезозое. Сальвинии – мелкие водные папоротники, замечательной чертой которой является разноспоровость. Мужские и женские гаметофиты развиваются из микро- и мегаспор, которые образуются в микро- и мегаспорангиях, заключенных в микро- или мегасорусах. Сорусы образуются на спороносных сегментах погруженного листа. Мегасорусы, обычно на ножках, закладываются первыми у основания сегмента. Микросорусы, как правило, сидячие и размещаются на остальной части сегмента. Все сорусы по форме и величине сходны между собой. Мегасорус состоит из нескольких (до 25) мегаспорангиев, расположенных или одиночно на короткой толстой ножке, или группами на ветвистых ножках. В мегаспорангиях развивается только одна крупная мегаспора. Она остается все время окруженной стенкой спорангия и опадает вместе с ней. Окружающая мегаспору пенистая масса играет роль плавательного приспособления как для мегаспоры, так и для развивающегося женского гаметофита.
В микросорусах сальвинии ушковидной образуется до 500 микроспорангиев. В каждом микроспорангии образуется 32 микроспоры. Микроспоры имеют ясно выраженный трехлучевой рубец. Созревшие микроспоры по 4 погружены в отвердевшую плазматическую массу, называемую массулой. Замечателен тот факт, что ископаемая сальвиния Рейза (Salvinia reusii) обладала амфиспорангиатными сорусами с многочисленными микроспорангиями и меньшим числом мегаспорангиев. У современных сальвиний были отмчены случаи развития наряду с нормальными не свойственных им амфиспорангиатных сорусов.
Сорусы созревают и погружаются на дно водоемов. У сальвинии плавающей (однолетнего растения) это происходит осенью. После сгнивания спорангия, поднимаются на поверхность. Микроспоры дают начало сильно редуцированным мужским гаметофитом, а мегаспоры прорастают в более крупные женские. Прорастание спор происходит внутри спорангия. Прорастающая микроспора прорывает стенку спрангия и образует вначале трехклеточный мужской гаметофит. При прорастании мегаспоры ее оболочка разрывается на три лопасти, и зеленый женский гаметофит выступает наружу в виде веера.
2.8. Отдел Голосеменные (Pinophyta)
Растения, относящиеся к отделу голосеменных (Pinophyta) отличаются от всех остальных представителей растительного мира тем, что производят семена. А все семенные растения, как голосеменные, так и цветковые, являются разноспоровыми. Это значит, что споры у них двух разных типов – микроспоры (мужские споры) и мегаспоры (женские споры). Первые дают начало мужскому гаметофиту, а вторые – женскому. В этом отношении семенные растения не отличаются от таких разноспоровых бесеменных растений, как селагинелла, полушник, марсилея или сальвиния. Тем не менее между ними имеются весьма существенные различия.
В то время как у огромного большинства бессеменных высших растений мегаспоры освобождаются из мегаспорангия, у семенных растений их единственная зрелая мегаспора остается постоянно заключенной внутри мегаспорангия и здесь же, внутри мегаспоры, происходит развитие женского гаметофита и процесс оплодотворения. Определенный шаг в этом направлении уже можно наблюдать у разноспоровых бессеменных растений. Так, у некоторых видов селагинеллы мегаспора остается внутри мегаспорангия и после оплодотворения, вплоть до значительного развития зародыша и даже молодого растеньица. Еще больше приближение к семенным растениям наблюдается у некоторых вымерших плауновидных, например у каменноугольных родов миадесмия (Miadesmia) и лепидокарпон (Lepidocarpon), у которых мегаспорангий был окружен покровом из свернувшегося спорофиллоида (или его боковых выростов). Это уже максимальное приближение к семени, но образование это является скорее аналогом, чем гомологом семени. Настоящее семя появляется только у голосеменных, при том значительно раньше, чем у таких растений, как миадесмия и лепидокарпон. В отличие от семенных плауновидных мегаспорангий у семенных растений окружен особым защитным покровом, называемым интегументом. Он умеет совершенно иное происхождение, чем покрывало мегаспорангиев миадесмии или лепитокарбона.
Мегаспорангий с окружающим его энтегументом называется семязачатком или семяпочкой. Это действительно зачаток семени, с которого после оплодотворения развивается семя. Развитие мегаспоры, также как
Пыльцевые зерна у всех саговниковых мелкие, широкоэллиптические, лодочковидные и имеют на дистальной стороне одну борозду.
Например, у рода эфедра (Ephedra), относящийся к классу Гнетовые (Gnetopsida) при прорастании микроспоры отделяется сначала первая проталлиальная клетка. При втором делении образуется ядро второй проталлиальной клетки, но самой клетки не образуется, так ядро это не отделяется перегородкой от ядра антеридиальной клетки. Далее ядро антеридиальной клетки делится, образуя ядро генеративной клетки. Оно образует два ядра, одно из которых дает начало двум спермиям. А цель пыльцевых зерен попасть на женский гаметофит. Когда это произойдет, образуется короткая пыльцевая трубка, которая через шейку архегония достигает яйцеклетки. Спермий, достигнув яйцеклетки, сливается с ней, образуя зародыш.
У представителей подкласса Хвойные (Pinidae) каждая функционирующая материнская клетка микроспор дает начало четырем гаплоидным микроспорам, которые в течение некоторого времени оказываются заключенными внутри оболочки микроспор. Число микроспор обычно очень велико, иногда исключительно большое. Они чрезвычайно легкие, что помогает их распространению ветром. Чрезвычайно своебразной является форма пыльцевых зерен. У большинства представителей семейства сосновых и подокарповых она бобовидная, с двумя выступами – воздушными мешками. Для большинства видов тсуги (Tsuga) типична шарообразная пыльца с одним опоясывающим воздушным мешком. Шарообразные безмешковые пыльцевые зерна имеются у лиственницы.
2.9. Отдел Цветковые (Magnoliphyta)
Цветковые растения представляют собой самый большой отдел растительного мира. От всех остальных отделов высших растений, за исключением голосеменных, цветковые отличаются образованием семени. Представители обоих этих отделов являются растениями семенными, в то время как все остальные отделы являются бессеменными (расселение у них происходит посредством спор, а не семян). От голосеменных же цветковые отличаются прежде всего тем, что семязачатки (семяпочки) у них заключены в более или менее замкнутую полость завязи (образованный одним или несколькими сросшимися плодолистиками). Благодаря тому что семязачатки заключены в полость завязи, пыльца попадает не непосредственно на микропиле семязачатка, а на рыльце. Наличие рыльца является характернейшей особенностью цветковых растений и фактически главным отличием цветка от стробилов голосеменных.
В связи с крайним упрощением гаметофитов цветковых растений и резким сокращением их онтогенеза они утратили гаметангии – антеридии и архегонии. Половое поколение цветковых растений лишилось половых органов. В результате формирования самих гамет, особенно мужских гамет, также ускоряется и они крайне упрощаются.
Эволюция в сторону сокращения стадий развития гаметофитов и ускорения их онтогенеза началась уже среди голосеменных, где некоторые хвойные, а также роды гнетум и вельвичия приблизились к цветковым растениям по степени редукции и строению мужского гаметофита, а гнетум и вельвичия также по лишенному архегониев женскому гаметофиту. Однако гаметофиты цветковых растений достигли максимально возможного упрощения и более ускоренного развития.
Параллельно с дифференциацией клеток стенки микроспорангия происходят события, ведущие к формированию микроспор. В результате ряда последовательных митотических делений из первичных спорогенных клеток образуются микроспороцисты, или материнские клетки микросор. Из каждого микроспороциста в результате меойтического деления образуется тетрада гаплоидных микроспор.
В период между окончанием мейоза и освобождением сестринских спор из каллозовой оболочки протекают процессы формирования спордермы (оболочки микроспор). В тетрадном периоде вокруг протопласта микроспоры прежде всего образуется новая оболочка – примэкзина. Вскоре после начала образования примэкзины начинают выявляться те области спородермы, где формируются специальные места, служащие для выхода пыльцевой трубки, называемые апертурами.
Сформировавшиеся микроспоры освобождаются из тетрады в результате быстрого растворения каллозовой оболочки. Высвободившиеся микроспоры быстро увеличиваются в размерах.
В развитии микроспор очень важную роль играют клетки тапетума. Накапливается всё больше данных, говорящих о том, что тапетум не только служит питательным материалом, но и принимает участие в формировании экзины.
Морфологическая активность цитоплазмы микроспоры заканчивается к началу деления её ядра. У большинства тропических растений ядро микроспоры начинает делится почти сразу же после окончания микроспорогенеза, но у растений умеренного и особенно холодного климата обычно наблюдается перерыв, которых длится от нескольких дней до нескольких недель.
Как известно, оболочка микроспоры состоит из двух главных слоев – экзины и интины. Строение этих двух слоев, также как и их толщина, является важным признаком характеризующим пыльцевые зерна разных растений. Интина представляет собой внутренний слой пыльцевого зерна. Она облегает его содержимое и служит материалом, образующим пыльцевую трубку. Экзина – это внешний слой оболочки пыльцевого зерна.
В экзине пыльцевого зерна имеются эластичные, гибкие, чаще тонкие места, служащие для выхода пыльцевой трубки, - апертуры. Апертуры бывают простые (борозды, щели, поры и др.) и сложные, у которых борозды, поры и прочие образования обладают дополнительной апертурой. Простые апертуры особенно характерны для пыльцы однодольных растений, а также для некоторых приметивных групп двудольных. У двудольных встречаются как сложные, так и простые апертуры.
Пыльца часто бывает собрана в диады, тетрады, поллиады, поллинии.
Семязачаток состоит из двух основных частей – мегоспорангия и окружающего его покрова, называемого интегументом. Интегумент не полностью закрывает мегаспорангий, на верхушке семязачатка его края не срастаются полностью и оставляют отверстие – микропиле.
В семязачатках наблюдаются два типа строения мегаспорангия и два типа семязачатков – крассинуцеллятные и тенуинуциллятные. Крассинуцеллятные семязачтки характеризуются относительно мощно развитым мегаспорангием, и поэтому материнская клетка мегаспор отделена от эпидермы мегаспорангия одним или несколькими слоями клеток. У тенуинуциллятных семязачатков, мегаспорангий которых более редуцирован, материнская клетка мегаспор лежит непосредственно под эпидермой мегаспорангия.
У цветковых растений в мегоспорангии образуется обычно одна материнская клетка мегоспор. Она претерпевает два деления мейоза, в результате чего аналогично появлению тетрады гаплоидных микроспор образуется тетрада гаплоидных мегаспор. Как и при формировании микроспор, появляются каллозовые оболочки, обеспечивающие изоляцию материнской клетки и мегаспор от окружающей клетки.
Мегаспоры тетрады бывают расположены по-разному, но чаще всего образуется линейная тетрада, реже – Т-образная и ещё реже – квадратная или тетраэдрическая. У громадного большинства цветковых растений возникают четыре мегаспоры, из которых нижняя и значительно реже верхняя увеличиваются в размерах и функционируют как мегаспора, а остальные три отмирают и разрушаются.
3. ВЫВОД
В своей курсовой работе я рассмотрела отделы высших растений, в частности те, которые относятся к разноспоровым: отделы Плауновидные (Lycopodiophyta), Хвощевидных (Equisetophyta), Папоротниковидные (Polypodiophyta), Голосеменные (Pinophyta или Gymnospermae), Цветковые (Magnoliophyta). В работе указала различие между семенными разноспоровыми высшими растениями, которые являются эволюционно более продвинутыми, и бессемеными разноспоровыми высшими растениями. Это различие заключается в возникновении семени. Также я подробно описала цикл размножения высших растений, без которого было бы неясно происхождение мегаспор и микроспор, обосновала факторы разноспоровости высших растений, выявила особенности разноспоровости.
4. СПИСОК ИСПОЛЬЗУЕМОЙ ЛИТЕРАТУРЫ
1. «Жизнь растений», том 4. – Москва. «Просвящение», 1978
2. «Жизнь растений», том 5(1).- Москва. «Просвящение», 1981
3. Большой биологический энциклопедический словарь.- Научное издательство «Большая Российская энциклопедия», Москва, 1999
4. Еленевский А.Г., Соловьева Н.П., Тихомиров В.Н. «Ботаника высших,или наземных растений». – Москва. Издательский центр «Академия», 2000