


Внутривидовая гибридизация хвойных пород
Лесная селекция и генетика – наука об улучшении качества и повышении продуктивности лесов путем управления наследственностью и изменчивостью древесных и кустарниковых растений. Она неразрывно связана с лесным семеноводством, задача которого заключается в получении сортовых семян древесных растений и районировании их пользования.
В программу интенсификации всех отраслей растениеводства, в том числе и лесных культур, наряду с агроприемами, включается использование улучшенного селекционно-генетическими методами посевного и посадочного материала. Таким образом, сорт лесных древесных пород так же, как и сорт в сельском хозяйстве, становится средством производства. Сор том называют совокупность растений с улучшенными по сравнению с исходными видами и сортами хозяйственными и биологическими признаками, устойчиво передающимися потомству при семенном и вегетативном размножении. Селекцией называют процесс выведения новых – и улучшения существующих сортов растений и пород животных. Слово селекция в переводе с латинского означает отбор или выбор. Отбор лучших форм из дикорастущих видов растений – наиболее древний метод селекции. Первоначальное понятие селекции соответствовало содержанию работ по выведению новых сортов путем отбора и размножения лучших форм. Современная селекция применяет отбор, используя при создании исходного материала гибридизацию, полиплоидию и мутагенез. Однако и в настоящее время отбор остается основой селекционной работы: с отбора исходных форм начинается выведение сорта, отбором заканчивается апробация форм и признание нового сорта.
Наука, изучающая закономерности наследственности и изменчивости, называется генетикой. Наследственность неразрывно связана с изменчивостью. Закономерности наследственности и изменчивости растений, установленные и разрабатываемые генетикой, составляют основу селекционной работы по выведению новых сортов. Генетика же заимствует и использует для обобщения и установления закономерностей наследственности и изменчивости фактический материал, получаемый селекцией в процессе создания сортов. Селекция растений имеет и собственные задачи по управлению процессами сортоводства, которые являются одной из форм направленной эволюции. Селекция – это учение об искусственном отборе в широком смысле этого слова. Изучение закономерностей естественного отбора позволило Ч. Дарвину селекцию из ремесла и искусства превратить в науку. Несмотря на выделение генетики в самостоятельную науку, селекция, как учение о закономерностях выведения сортов на организменном и на популяционном уровне, не лишилась самостоятельности, а приобрела новый более совершенный уровень своего развития.
Селекционно-генетические и семеноводческие знания о лесных древесных и кустарниковых растениях входят в состав смежной науки – лесной селекции и генетики. Задача этой науки заключается в разработке селекционно-генетических методов повышения продуктивности и улучшения качества создаваемых лесов. Выполнение этой важнейшей задачи практической селекции в области лесоводства неотделимо от сортоводства древесных пород.
По способам размножения сорта древесных растений можно разделить на четыре группы: сорта-клоны, сорта-линии, сорта-популяции и сорта гибридного происхождения, или сорта-гибриды.
Сорта, получаемые скрещиванием особей с разной наследственностью и отбором из гибридных семей, называются гибридными. Эти сорта преобладают в настоящее время во всех областях растениеводства, так как они обладают выделяющимися качествами вследствие последовательного накопления на различных ступенях сложного скрещивания важнейших свойств, которыми должен обладать сорт.
В лесном хозяйстве и декоративном садоводстве использование диких видов и форм в культуре приобретает в настоящее время особое значение Возможность применения селекции к лесным древесным растениям не вызывает ни у кого сомнений, а переход при ведении лесного хозяйства от рода и вида к сорту – реальная практическая необходимость.
1. Селекция хвойных пород
1.1 Народнохозяйственное значение и селекция хвойных пород
Хвойные – основные породы-лесообразователи Советского Союза. Они занимают большие площади таежной зоны Евразии, составляя основу лесосырьевой базы страны. Хвойные леса СССР оказывают существенное влияние на климат Северного полушария планеты. Основное хозяйственное значение у хвойных имеет древесина, которая широко используется для изготовления пиломатериалов, в качестве сырья для целлюлозно-бумажной промышленности, для получения искусственного шелка, производства музыкальных инструментов, токарных изделий и т.д.
Древесина сосны с красноватым ядром и желтовато-белой заболонью с четкими границами годичных колец издавна пользуется большим спросом. Круглый лес (столбы, бревна, рудничная стойка), пиломатериалы (балки, брусья, доски, тес), колотый лес и щепные товары (клепка, щепа) – основные сортименты сосновой древесины. Мягкая ядровая древесина кедра сибирского используется для производства карандашей. Древесина лиственницы, очень стойкая в воде и противостоящая гниению, успешно используется для гидротехнических сооружений, подводных и мостовых брусьев, рудничной стойки и др. С красноватым ядром и желтовато-белой заболонью древесина лиственницы очень Декоративна, применяется дли отделки Интерьеров общественных зданий. Красоту рисунка древесины лиственницы дополняют четко выраженные годичные слои. Древесина богата смоляными ходами. Древесина ели и пихты без ядра. Она применяется как строевой лес, и как сырье для целлюлозно-бумажной промышленности, для получения искусственного шелка, и изготовления музыкальных инструментов. Древесина можжевельника отличается прочностью, не имеет смоляных ходов и обладает высокой декоративностью благодаря коричневатому ядру и узкой беловатой заболони. Годичные слои у нее извилистые, поздняя древесина их резко отличается от ранней. Древесина можжевельника пользуется большим спросом для изготовления токарных изделий, тростей, шахмат, игрушек и сувениров.
Для обширных областей Европы, Азии, Северной Америки и отчасти Африки древесина хвойных издавна была основным строительным материалом, из которого возводились жилища, хозяйственные постройки, культурные и общественные здания. Значение древесины хвойных как идеального по своим техническим качествам столярного и строительного материала особенно возросло за последние два столетия, когда механизация производства потребовала однородного сырья древесины. Однако наибольшее значение приобрела древесина хвойных в целлюлозно-бумажной промышленности, где ценится длина волокна. Трахеиды, из которых главным образом сложена древесина хвойных, оказались самым длинноволокнистым элементом среди современных древесных растений. Наибольшей длиной из хвойных северного полушария отличаются трахеиды ели (4300 мкм) и лжетсуги (4500 мкм).
Хвойные широко используются для добычи смолы, дубильных веществ и другого ценного сырья для промышленности. При подсочке хвойных добывают живицу для производства скипидара и канифоли, в коре ели и лиственницы содержатся дубильные вещества, из коры можжевельника добывают скипидар для лаков, а смола коры пихты содержит до 25% скипидара. Семена кедровых сосен и шишки можжевельников используются в питании человека. Из хвои пихты, ели, сосны и можжевельника получают эфирные масла и витамин С, используемые в медицине.
Продолжительная и интенсивная эксплуатация хвойных поставила человечество перед необходимостью забот о сохранении и селекции их на повышение продуктивности и качества древесины, живицы и др. Дальнейшая интенсификация лесного хозяйства предполагается не только за счет селекции местных хвойных видов, форм и сортов, но и широкой интродукции хозяйственно ценных зарубежных пород.
1.2 Систематика и распространение хвойных пород
Современные хвойные, или пиниды, насчитывают 5 порядков, 7 семейств, 57 родов и 560 видов.
Среди современных хвойных самыми древними семействами являются араукариевые, подокарповые и сосновые. Пыльцевой анализ показал, что предки современных представителей хвойных существовали еще до мелового периода, т.е. до появления цветковых растений. За время своего существования хвойные подверглись значительной эволюции. Однако хвойные отличаются заметной стабильностью генетической организации. Эволюционная стабильность хвойных объясняется общей консервативностью возникших в ходе эволюции хвойных морфологических структур. Говоря о темпах хромосомной эволюции, генетики отмечают, что у сосновых они в десятки и сотни раз ниже по сравнению с другими организмами. Вместе с тем скорость эволюции структурных генов, ответственных за синтез белков у сосновых, соответствует средней скорости эволюции других организмов.
Долговечность хвойных (от 300 до 2000–3000 лет) определяется особенностями строения стволов, отличающихся от стволов других голосеменных более развитой древесиной и менее развитыми кроной и сердцевиной. Ксилема хвойных по объему на 90–95% состоит из трахеид, длина которых у разных видов сильно варьирует (от 0,5 до 11 мм). В процессе эволюции длина трахеид постепенно уменьшается. Число пор на каждой трахеиде от 30 до 50 шт., они сосредоточены на концах трахеид, где интенсивность тока пасоки наибольшая. Трахеиды с округлыми окаймленными порами – достаточно совершенные водопроводящие элементы.
Древесной паренхимы у хвойных очень мало, у тисовых и араукариевых она отсутствует. Сердцевинные лучи узкие, однорядные или двурядные (реже более широкие) от 1 до 60 клеток по высоте.
В древесине и коре современных хвойных много смоляных ходов. Они имеют форму длинных узких каналов, наполненных эфирными маслами, смолой и бальзамами, выделяемыми выстилающими клетками. Только для представителей семейства сосновых характерны постоянные нормальные (не травматические) смоляные ходы во вторичной древесине. У огромного большинства хвойных, не имеющих смоляных ходов в древесине в норме, они образуются как реакция на поранение ствола. Однако у араукариевых смоляные ходы не образуются даже при повреждениях стволов. Смола у них откладывается в трахеидах, окаймляющих сердцевинные лучи.
Хвойные – разнополые, однодомные, реже двудомные растения. Микростробил, который часто называют мужским колоском, состоит из укороченной оси, на которой расположены сильно редуцированные микроспорофиллы с микроспорангиями. В систематики и филогении хвойных особый интерес представляет строение женских стробилов, которые называют макро- или мегастробилами. Уменьшение числа макростробил – общее направление эволюции хвойных. У тисовых в результате далеко зашедшей редукции числа макростробилов формируются одиночные, односеменные макростробилы. Эволюция большинства хвойных шла в сторону возрастания компактности собрания макростробилов, уменьшение же числа частей играло второстепенную роль. В итоге возникли компактные шишки – типа шишек сосны, ели, араукарии и других хвойных.
У большинства представителей семейства сосновых (сосны, ели, пихты) пыльца бобовидной формы с двумя выступами или воздушными мешками. Для видов тсуги типична шарообразная пыльца с одним опоясывающим воздушным мешком. У лиственницы и ложнотсуги пыльцевых мешков нет и пыльца имеет шарообразную форму.
Ветроопыление у хвойных, т.е. перенос пыльцы на семяпочку движением воздуха, происходит различно. Наиболее известный тип опыления характерен для сосны, имеющей пыльцу с двумя воздушными мешками. Семенные чешуйки в шишке сосны в период цветения несколько раздвигаются, что свидетельствует о готовности семяпочек к принятию пыльцы. Обильное количество пыльцы, доставляемое ветром, попадает в промежутки между семенными чешуями и задерживается так называемыми рыльцевыми расширениями края микропиле (пыльцевхода). У сосны эти расширения узкие, у ели они шире. К этому времени (а иногда и раньше) микропиле начинает выделять так называемую опылительную жидкость, выступающую на его верхушке в виде капли (опылительная капля). Пыльцевые зерна легко смачиваются опылительной жидкостью и быстро погружаются в нее. В результате пыльцевое зерно как бы засасывается внутрь и достигает верхушки мегаспорангия (нуцеллуса), где и прорастает. Благодаря воздушным мешкам, играющим роль своеобразных плавательных приспособлений, пыльцевое зерно по мере продвижения вверх по микропилярному каналу все время остается ориентированным дистальным полюсом вниз по направлению в пыльцевую камеру, образованную краями нуцеллуса. В таком положении пыльцевое зерно легко прорастает и образует пыльцевую трубку. После опыления чешуи сближаются и остаются плотно прижатыми друг к другу до созревания семян.
Развитие зародыша семени у хвойных начинается с образования предзародыша, или зачаточного зародыша (проэмбрио). После этого начинается развитие собственно зародыша. Увеличиваясь, подвесок не только проталкивает зародыш вглубь ткани гаметофита и служит для них гаусториальным органом, но и является, вероятно, секреторным органом, разрушающим с помощью ферментов клетки эндосперма.
Семена хвойных очень разнообразны по величине и форме. Крупные семена с деревянной кожурой бескрылые, а кожистые и перепончатые большинства хвойных снабжены одним крыловидным придатком или двумя-тремя небольшими крыльями, что способствует их распространению ветром. Сочный, ярко окрашенный покров на семенах ряда хвойных способствует их распространению животными.
Исчезновение воздушных мешков у пыльцы лиственницы и, лжетсуги связано с отсутствием опылительной жидкости. Следует, однако, отметить, что у пихты и кедра опылительная жидкость не выделяется, но пыльцевые зерна снабжены воздушыми мешками, хотя у кедра наблюдается тенденция к их редукции. У пихты микропилярная область превращается в косую рыльцевую воронку, улавливающую пыльцу. Пыльцевые зерна не достигают нуцеллуса. Вместо этого нуцеллус растет кверху, и этим достигается контакт с пыльцевыми зернами. У кедра, как и у пихты, в результате согласованных изменений микропиле и нуцеллуса пыльцевые зерна оказывая на нуцеллусе, где и прорастают.
Оплодотворение у хвойных, за исключением сосны, происходит вскоре после опыления. У видов сосны между опылением и оплодотворением проходит от 12 до 14 мес.
В процессе длительной эволюции ряд видов хвойных успешно приспособился к изменяющимся условиям среды благодаря совершенствованию и закреплению в потомстве древних генетических структур, определяющих соответственно проявление конкурентоспособных свойств. Подтверждением этого могут служить таежные леса с преобладанием хвойных пород, занимающие огромные территории холодной зоны в Северном полушарии, образующие чистые и смешанные хвойно-лиственные древостои. Ксероморфность ассимиляционного аппарата, Относительная долговечность, выработка защитных средств (живицы, эфирных масел), строение водопроводящей системы, ветроопыляемость и ряд других особенностей строения обеспечили хвойным одно из ведущих положений в образовании современных лесных формаций на земном шаре. Знание особенностей исторического развития, или филогении, видов позволяет выработать методы управления закономерностями наследственности и изменчивости при их искусственном разведении.
1.3 Селекционные методы улучшения хвойных пород
Селекционно-генетическое улучшение лесообразующих хвойных пород в нашей стране осуществляется в плановом порядке. Селекционные программы составляются по следующим направлениям: секция на быстроту роста и продуктивности сосны, ели, лиственницы, пихты, можжевельника; селекция на смолопродуктивность сосны; селекция на урожайность кедровых сосен; селекция на резонансность древесины ели; селекция на декоративность и устойчивость к промышленным выбросам сосны, ели, лиственницы, пихты, можжевельника, псевдотсуги, тсуги и др. Комплексное использование массового, семейного и клонового отборов – основной метод современной селекции хвойных.
Селекционная практика показала, что отбор целесообразно вести по небольшому числу признаков. Успех работы по селекции, в том числе и хвойных пород, во многом зависит от методики изучения прямых селекционных признаков и корреляции их с удобными для использования косвенными признаками. Генетическая обусловленность признака у лесообразующих хвойных определяется путем математического разложения общей фенотипической изменчивости на генотипическую и паратипическую (средовую) изменчивость. Наследуемость в общем смысле, без смены поколений в природных популяциях хвойных, изучается дисперсионным анализом и другими математическими методами при отборе на общую комбинативную способность. Для определения наследуемости в узком смысле слова требуется изучение характера наследования признака (доминирование, эпистаз или аддитивное наследование), которое используется при отборе на специфическую комбинативную способность. Эти знания необходимы для практической работы по селекции, так как они позволяют раскрыть генетическую характеристику селекционного признака.
Селекционно-генетическая программа улучшения хвойных включает такие вопросы:
1. Постановка цели и задачи. Современное состояние вопроса и перспективы на обозримое будущее (5, 10–20, 50 и более лет).
2. Исходный материал для селекции (родовой комплекс лесообразователей местных пород и интродуцентов).
3. Методы отбора и интенсивность ведения хозяйства (массовый и групповой отбор с определением общей комбинативной способности в диких популяциях.
4. Отбор на общую и специфическую комбинативную способность, индивидуальный отбор и плантационное хозяйство.
5. Создание постоянной семеноводческой базы для получения сортовых семян в соответствии с направлением ведения хозяйства.
6. Организация выращивания сортового посевного и посадочного материала для лесных культур и промышленных плантаций.
7. Закладка культур и промышленных плантаций по выращиванию заданных сортиментов древесины.
8. Разработка селекционных методов рубок ухода, стимулирующих рост деревьев в искусственных посадках.
Таким образом, селекционные методы улучшения хвойных органически входят в общую систему лесоводческих мероприятий по интенсификации лесохозяйственного производства. Особое значение в деле повышения производительности хвойных лесов нашей страны приобретает создание популяционного сортоводства на основе системного генетического анализа естественных насаждений. Поэтому знания частной генетики хвойных в целом, а также отдельных родовых комплексов и видов необходимы лесоводам, чтобы успешно осуществлять селекционно-семеноводческую программу в лесах нашей страны.
1.4 Исходный материал для селекции
Селекционная работа начинается с подбора исходного материала в соответствии с избранным направлением и поставленной целью. Исходным материалом в селекции называют культурные и дикие формы растений и популяций, используемые для выведения новых сортов. В современной селекции в качестве исходного материала используются несколько форм и популяций.
Естественные (природные) популяции – дикорастущие формы, местные сорта культурных растений и образцы мировой коллекции сельскохозяйственных растений Всесоюзного института растениеводства имени Н.И. Вавилова.
Гибридные популяции – внутривидовые популяции, создаваемые в результате скрещивания сортов и форм в пределах одного вида, и популяции, полученные от скрещивания особей разных видов и родов растений (межвидовые и межродовые).
У перекрестноопыляющихся растений одним из новых источников материала являются самоопыляемые линии, или инцухт-линии. Их получают многократным принудительным самоопылением перекрестноопыляющихся растений. Лучшие линии скрещивают между собой, получают межлинейные гибриды или их скрещивают с другими сортами для создания гетерозисных гибридов. В результате скрещивания получают гибридные семена, которые используют в течение 1 года. Сорта-гибриды этих межлинейных скрещиваний в отличие от сортов гибридного происхождения нужно ежегодно воспроизводить.
Искусственные мутации и полиплоидные формы создаются путем воздействия на растения различными мутагенами (радиацией, химическими веществами, температурой, прививками и др.).
Ценным источником сортов древесных растений являются формовые разнообразия естественных популяций и интродукции новых видов из других флористических зон. Введение в культуру новых форм и сортов в будущем улучшит качество наших лесов. Селекция поможет создавать сортовые промышленные плантации в соответствии с ростом потребности в определенных сортиментах древесины и других полезностях леса.
В процессе развития человечества наши леса, как и остальные компоненты природы, подвергаются значительным изменениям. Так, в течение многих столетии выборочные рубки деревьев лучшего качества без заботы о восстановлении лесов отрицательно повлияли на лесонасаждения. Это сокращает мировой лесной генофонд лесных древесных популяций.
Работа по селекции всех видов растений, в том числе и древесных пород, основывается на использовании естественного генофонда местных древесных пород лесообразователей и интродуцентов. Сохранение генофонда древесных растений как для осуществления мероприятий по улучшению свойств древесных растении в настоящее время, так и для селекционной работы и составлении перспективного ассортимента в будущем становится в центре внимания лесной селекции. Учет и исследование генофонда – основа каждой селекционно-генетической программы.
Все особи вида имеют общие характерные черты, а также свои индивидуальные генотипические признаки, что в целом отражает генетическое разнообразие вида, которое называют генофондом вида.
Генофонд древесных растений обследуется в настоящее время путем изучения генетической структуры популяций основных лесообразователей. При выборе признаков, фенотипическое проявление которых могло бы дать информацию о генофонде популяции, большое значение имеют два показателя: генетический и экономический. Генетическая характеристика генофонда включает степень наследуемости признаков, выражающую сущность фенотипической ценности популяции. Анализ генофонда популяции возможен именно по фенотипическим выражениям тех признаков, которые характеризуются высокой степенью наследуемости. В свою очередь степень наследуемости определяется по регрессии «родители – потомки», анализу сибсов и полусибсов и в некоторых случаях проверкой коэффициента наследуемости аналогичной изменчивости эталонным методом в географических клоновых культурах с выявлением соотношения между генетической и фенотипической дисперсиями.
Экономическая характеристика генофонда имеет важное значение при изучении генофонда по хозяйственно ценным признакам. Но при этом не следует забывать, что пока еще невозможно уверенно прогнозировать экономическую ценность многих признаков для будущего. Это во многом зависит от уровня развития промышленности по использованию и переработке древесины и других полезностей лесных древесных растений в будущем. В связи с этим необходима разработка мероприятий по сохранению в наших лесах всей полноты генофонда. Поэтому селекционные размеры популяций должны быть большими, что возможно при сохранении значительного числа особей каждой популяции.
2. Гибридизация хвойных пород
2.1 Гибридизация
Скрещивание особей, отличающихся друг от друга хотя бы одним аллелем, называется гибридизацией. Гибридами соответственно называют потомство, полученное в результате искусственного или свободного (спонтанного) скрещивания особей с разной наследственностью. Различают внутривидовую, межродовую и межсемейственную гибридизацию. Скрещивание особей различных форм и сортов, принадлежащих к одному виду, называется внутривидовой гибридизацией. Скрещивание особей, принадлежащих к разным видам одного рода, разным родам и разным семействам, называется отдаленной межвидовой, межродовой и межсемейственной гибридизацией.
Гибридизацию нельзя рассматривать как простое арифметическое суммирование признаков и свойств растений. Родительские организмы передают потомству не признаки, а гены, на основе которых в каждом поколении гибридов признаки, контролируемые этими генами, развиваются вновь. Гибридизация используется в качестве способа изучения наследования, получившего название гибридологического метода генетического анализа. Этот метод генетики основан на принципе менделеевского анализа наследования и взаимодействия отдельных генов у организмов. При этом у гибридного потомства изучается наследование не совокупности признаков, а одного, двух или трех контрастных признаков в ряду последовательных поколений с применением индивидуального анализа потомства от каждого гибридного растения.
2.1.1 Гибридизация как метод селекции
Гибридизация как метод селекции включает комплекс приемов, направленных на получение гибридных растений с изменением наследственности и использованием ее для выведения новых сортов. Создавая гибридизацией нужный исходный материал, удается значительно ускорить ход селекционного процесса. Последовательным скрещиванием наследственно расщепляющихся родительских форм селекционеры создают новые формы растений. Выведение новых сортов, в которых с помощью гибридизации достигается сочетание хозяйственно ценных свойств большого количества родительских форм названо синтетической селекцией. Гибридизацию относят к категории комбинативной селекции, так как основной целью при этом является получение потомства с новой совокупностью генетически обусловленных признаков и свойств. Последующим отбором и направленным воспитанием гибридного потомства новые ценные признаки и свойства закрепляются и усиливаются.
2.1.2 Содержание метода гибридизации
Содержание и порядок работы по селекции методом гибридизации:
1. Цель работы и разработка модели (образа) будущего гибридного сорта.
2. Изучение генетического потенциала (наследственности) исходного материала.
3. Подбор родительских пар.
4. Подбор и хранение пыльцы.
5. Подготовка женских цветков к опылению (кастрация и изоляция).
6. Проведение опыления (техника скрещивания).
7. Наблюдение за развитием гибридных семян и уход за материнскими растениями.
8. Сбор гибридных семян и выращивание гибридного потомства.
9. Отбор лучших гибридных форм и выделение из них отдельных растений (кандидатов в сорта) для сравнительного испытания на сортоиспытательных участках.
10. Разработка методов массового размножения нового сорта для производства.
Сортам, полученным в итоге гибридизации, присваивается название, которое включает название исходных сортов или видов. Очень часто селекционеры присваивают сортам произвольные названия, подчеркивая их характерные сортовые особенности. Например, лесными селекционерами СССР получены региональные зимостойкие сорта белого пирамидального тополя от скрещивания местных форм белого тополя с тополем Боллеана. Этим сортам даны соответствующие названия: Советский пирамидальный селекции А.С. Яблокова, Уральский пирамидальный селекции Н.Н. Коновалова и т.д.
2.2 Типы скрещиваний, применяемые при гибридизации
Изменение наследственности исходного материала для селекции при гибризации осуществляется однократными, или простыми, скрещиваниями и многократными скрещиваниями, названными сложными или ступенчатыми скрещиваниями. Наиболее распространенное деление скрещиваний на простые и сложные с последующим подразделением на целый ряд типов можно представить такой схемой.
Простыми скрещиваниями называют однократные скрещивания между двумя родительскими формами. Если родительские виды или сорта обозначить буквами, то этот тип скрещивания можно изобразить как АЧБ или ВЧГ и т, д., после которых в гибридном потомстве проводится отбор элитных растений и оценка их потомства. При простых скрещиваниях гибриды получаются на основе комбинаций генов материнской и отцовской форм. Простые парные скрещивания имеют большее значение при внутривидовой гибридизации.
Диаллельные скрещивания – каждая испытываемая линия, форма или сорт скрещивается со всеми другими линиями или сортами во всех возможных комбинациях. Например, АЧБ, АЧВ, АЧГ, АЧД, АЧЕ и т.д. Число всех возможных комбинаций при диаллельных скрещиваниях может быть очень большим и будет возрастать – по мере увеличения количества исходных линий, форм или сортов. Так, при диаллельном скрещивании 10 линий число всех возможных комбинаций составляет 45, а при 100 линиях уже 4950. Поэтому на практике диаллельные скрещивания обычно проводят в пределах сравнительно небольших групп, в которых бывает примерно по 10 линий или сортов. Диаллельные скрещивания можно применять в работе по гибридизации с древесными растениями с целью изучения варьирования признаков в гибридном потомстве, а также определения отобранных по фенотипу деревьев на проявление хозяйственно ценного признака в гибридном потомстве.
![]()
![]()
![]()
![]()
![]()

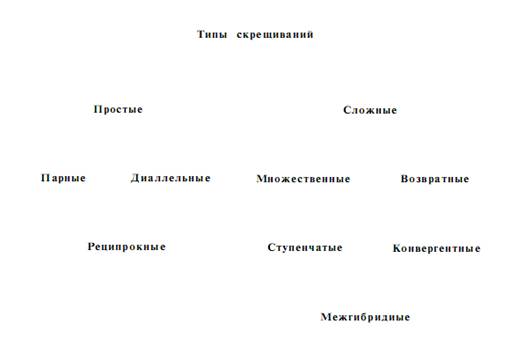
Реципрокными скрещиваниями называются скрещивания растений, при которых каждый из двух сортов или видов в одном случае является материнской формой, во втором – отцовской. Например, скрещивание, осуществленное по схеме АЧБ и БЧА. Первое скрещивание АЧБ называют прямым, второе БЧА –обратным. Реципрокные скрещивания часто используют с разведывательной целью, чтобы выяснить какую из двух форм лучше взять в качестве материнской, а какую в качестве отцовской. Реципрокные скрещивания важны при отдаленной гибридизации, так как часто успех работы решается удачным подбором отцовских и материнских видов. Для реципрокных скрещиваний составляют план, или сетку, скрещиваний.
Сложными скрещиваниями называются скрещивания, когда в гибридизацию вовлекается более двух родительских форм или когда гибридное потомство повторно скрещивается с одним из родителей. Сложные скрещивания в селекционной практике имеют значительно большее значение, чем простые.
Множественные скрещивания, или поликросы, – это такие скрещивания, когда материнское растение опыляется смесью пыльцы нескольких видов и сортов. Этот вид скрещивания схематически можно изобразить так: АЧ (Б+В+Г+Д и т.д.), где буквой А обозначен материнский сорт, а буквами Б, В, Г, Д – отцовские сорта, от которых берется пыльца для составления смеси. Множественные скрещивания осуществляются двумя способами: искусственным опылением материнского растения смесью пыльцы нескольких отцовских форм: свободным опылением материнского растения с помощью ветра или насекомых, когда материнские и отцовские растения высаживаются рядом на одной и той же семенной площадке. Метод множественного опыления успешно применил к плодовым культурам И.В. Мичурин. В селекции многих кормовых многолетних трав и некоторых древесных растений сейчас широко используется этот метод, получивший название метода поликроссной селекции.
Основные условия успешного применения метода множественных скрещиваний: растения должны быть многолетними, самостерильными, обладать способностью к клонированию и иметь одинаковые сроки цветения. Таким условиям могут удовлетворять многие древесные породы.
Возвратные скрещивания, или беккроссы, – скрещивания, при которых гибрид повторно скрещивается с одной из родительских форм. В природных условиях повторные скрещивания спонтанных гибридов с одной из родительских форм называются интрогрессивной гибридизацией. В практике селекционной работы беккроссы называют повторными (возвратными) скрещиваниями. Возвратные скрещивания можно записать схематически так: 1-й год – АЧБ; 2-й год – (АЧБ) ЧА или (АЧБ)ЧБ. Этот тип скрещивания широко применяется в селекционной практике. Он используется в тех случаях, когда у ценных по комплексу признаков сортов имеется дефект, который желательно устранить. Тогда новый сорт стал бы совершеннее, расширились бы возможности его практического использования. При возвратных скрещиваниях тот сорт, от которого хотят взять основной комплекс признаков, берется при первом скрещивании обычно в качестве материнского, а при повторных скрещиваниях он используется в качестве отцовского. Изменчивость гибридного потомства при возвратных скрещиваниях суживается. Полученные гибриды дают расщепление в отношении 1:1 (1-й год – ааЧАА–Аа; 2-й год – АаЧаа-1АаЧ1 аа). Селекционная цель достигается сравнительно быстро.
Насыщающие и конвергентные скрещивания – повторные возвратные скрещивания. Этот метод часто применяется при выведении сортов устойчивых к болезням. При насыщающих скрещиваниях признаки и свойства одного из родителей почти полностью вытесняются за исключением немногих генов. Во многих случаях это бывает нежелательным. Во избежание этого разработана система конвергентных скрещиваний. Конвергентные скрещивания представляют собой дальнейшее развитие метода возвратных скрещиваний. Метод заключается в том, что после получения F1 дальнейшее скрещивание проводят в двух направлениях. В одном случае гибриды повторно скрещивают с материнским сортом, а во втором – с отцовским. В результате получают две сближенные линии. Их скрещивают между собой и среди гибридного потомства производят отбор. После возвратных скрещиваний и сближения линий гибридное потомство проявляет менее сложный характер расщепления. Вследствие этого среди потомства легче найти желаемую комбинацию признаков.
Ступенчатые скрещивания – полученный от простого скрещивания гибрид повторно скрещивается не с родительской формой, а с третьим сортом или видом растений, затем с четвертым и т.д. Таким образом, в этих скрещиваниях участвуют несколько родительских форм, которые последовательно или ступенчато включаются в гибридизацию. При ступенчатых скрещиваниях создается гибридный материал, включающий наследственные свойства нескольких сортов, или видов растений.
Межгибридными скрещиваниями называют такие, при которых объединение наследственности нескольких родителей осуществляют не последовательно, как при ступенчатой гибридизации, а параллельно после предварительного получения простых гибридов и последующего их скрещивания.
Межгибридные скрещивания–главный метод при создании гетерозисных гибридных семян кукурузы и некоторых других культур.
Все типы скрещиваний успешно применяются к лесным древесным растениям. Они позволяют в ряде случаев создавать оригинальные комплексные программы применительно к особенностям той или иной группы видов. Перспективным приемом создания таких программ является включение внутривидовой и межвидовой гибридизации по родовым комплексам.
Лесные древесные растения выращиваются обычно с целью получения вегетативной массы, поэтому стерильность гибридных растений не может препятствовать их выращиванию в промышленных масштабах. В современной лесной генетике и селекции гибридизация играет более важную роль, чем в селекции сельскохозяйственных культур. Применение межвидовой гибридизации близких видов в некоторых родовых комплексах древесных пород из разных районов произрастания имеет большое значение для получе