Эволюция энергетических процессов у эубактерий
В главах 11 и 12 были обсуждены проблемы возникновения первичной клетки из гипотетической протоклетки и последующего пути прогрессивной эволюции первичной клетки. Как было обнаружено в 70-х гг., на раннем этапе этого пути могло произойти выделение трех основных ветвей, каждая из которых самостоятельно и независимо эволюционировала, результатом чего явилось существование в рамках прокариотной клеточной организации двух крупных таксономических групп: эубактерий и архебактерий.
Подавляющее большинство известных прокариот относится к эубактериям. Именно у них достаточно хорошо изучены основные этапы прогрессивной эволюции, связанные с совершенствованием энергетических процессов. Архебактерии представляют собой к настоящему времени разрозненные группы со специфическими типами энергетического метаболизма. Имеющегося материала явно не достаточно для создания каких-либо эволюционных построений внутри группы архебактерий.
ГЛАВА 13. БРОЖЕНИЕ. ТИПЫ ЖИЗНИ, ОСНОВАННЫЕ НА СУБСТРАТНОМ ФОСФОРИЛИРОВАНИИ
Наиболее примитивным способом получения энергии, присущим определенным группам эубактерий, являются процессы брожения.
ОБЩАЯ ХАРАКТЕРИСТИКА ПРОЦЕСCOВ БРОЖЕНИЯ
"Брожение" — это сугубо микробиологический термин. Он характеризует энергетическую сторону способа существования нескольких групп эубактерий, при котором они осуществляют в анаэробных условиях окислительно-восстановительные превращения органических соединений, сопровождающиеся выходом энергии, которую эти организмы используют. Поскольку брожение протекает без участия молекулярного кислорода, все окислительно-восстановительные превращения субстрата происходят за счет его "внутренних" возможностей. Процесс брожения связан с такими перестройками органических молекул субстрата, в результате которых на окислительных этапах процесса высвобождается часть свободной энергии, заключенной в молекуле субстрата, и происходит ее запасание в молекулах АТФ. В процессе брожения, как правило, происходит расщепление углеродного скелета молекулы субстрата.
Круг органических соединений, которые могут сбраживаться, довольно широк. Это углеводы, спирты, органические кислоты, аминокислоты, пурины, пиримидины. Химическое вещество может быть подвергнуто сбраживанию, если оно содержит неполностью окисленные (или восстановленные) углеродные атомы. В этом случае есть возможность для окислительно-восстановительных преобразований между молекулами (или внутри одного вида молекул), возникающими из субстрата. В результате одна часть продуктов брожения будет более восстановленной, другая — более окисленной по сравнению с субстратом. Продуктами брожений являются различные органические кислоты (молочная, масляная, уксусная, муравьиная), спирты (этиловый, бутиловый, пропиловый), ацетон, а также CO2 и H2. Обычно в процессе брожения образуется несколько продуктов. В зависимости от того, какой основной продукт накапливается в среде, различают молочнокислое, спиртовое, маслянокислое, пропионовокислое и другие виды брожений.
Следовательно, в каждом виде брожения можно выделить две стороны: окислительную и восстановительную. Процессы окисления сводятся к отрыву электронов от определенных метаболитов с помощью специфических ферментов (дегидрогеназ) и акцептированию их другими молекулами, образующимися из сбраживаемого субстрата, т. е. в процессе брожения происходит окисление анаэробного типа.
Энергетическая сторона
Собственно энергетической стороной процессов брожения является их окислительная часть, поскольку реакции, ведущие к выделению энергии, — это реакции окисления. Существует несколько исключений из этого правила: некоторые анаэробы часть энергии при сбраживании субстрата получают также в результате его расщепления. Примитивность процессов брожения заключается в том, что из субстрата в результате его анаэробного преобразования извлекается лишь незначительная доля той химической энергии, которая в нем содержится. Продукты, образующиеся в процессе брожения, все еще содержат в себе значительное количество энергии, заключавшейся в исходном субстрате. Чтобы четче представить разницу в энергетическом выходе процессов брожения и дыхания, приведем данные по изменению уровней стандартной свободной энергии для процессов гомоферментативного молочнокислого брожения и дыхания при одинаковом исходном энергетическом субстрате (глюкоза):
|
|
DG0' = –196,65 кДж/моль; | DG0' = –2870,22 кДж/моль. |
В процессе гомоферментативного молочнокислого брожения синтезируются 2 молекулы АТФ на 1 молекулу сброженной глюкозы; в процессе дыхания при полном окислении молекулы глюкозы образуется 38 молекул АТФ. В обоих случаях эффективность запасания выделяющейся энергии в макроэргических связях АТФ приблизительно одинакова.
При брожении некоторые реакции на пути анаэробного преобразования субстрата связаны с наиболее примитивным типом фосфорилирования — субстратным фосфорилированием. К синтезу АТФ по механизму субстратного фосфорилирования ведут катаболические реакции, которые в зависимости от своей химической природы могут быть разделены на два типа. Большинство относится к окислительно-восстановительным реакциям. Богатые энергией соединения возникают в процессе брожения на этапах анаэробного окисления. Например, окисление фосфоглицеринового альдегида (ФГА), катализируемое ФГА-дегидрогеназой, приводит к образованию богатого энергией метаболита — 1,3-дифосфоглицериновой кислоты (1, 3-ФГК). Анаэробное окисление пировиноградной или a-кетоглутаровой кислот приводит к образованию высокоэнергети-ческих метаболитов — ацетил-КоА40 или сукцинил-КоА соответственно.
40 Символ "КоА" обозначает кофермент А, связанный с ацильной группой (R-CO~S-KoA), "КоА-SH", находящийся в свободном состоянии.
Второй тип реакций связан с расщеплением субстратов или промежуточных продуктов, образующихся из них. Катализируются эти реакции ферментами, относящимися к классу лиаз. Например, у гетероферментативных молочнокислых бактерий высокоэнергетический ацетилфосфат образуется из ксилулозо-5-фосфата в реакции, катализируемой фосфокетолазой:
ксилулозо-5-фосфат + ФН ® ФГА+ ацетилфосфат + H2O.
К реакциям подобного типа относится также расщепление цитруллина, приводящее к синтезу карбамоилфосфата — соединения с макроэргической фосфатной связью:
цитруллин + ФН ® карбамоилфосфат + орнитин.
Богатые энергией соединения, образующиеся в реакциях рассмотренных выше типов, представляют в большинстве случаев ангидриды фосфорной кислоты или тиоэфиры органических кислот. Последние используются для синтеза АТФ через ферментативную стадию образования соответствующих ацилфосфатов:
ацил-КоА + ФН ® ацилфосфат + КоА-SH.
Из других высокоэнергетических соединений важное место в энергетике процессов брожения принадлежит фосфоенолпировиноградной кислоте (ФЕП). Эти соединения характеризуются тем, что свободная энергия, освобождающаяся при их гидролизе, находится в области значений от -35 до -88 кДж/моль и с помощью соответствующих ферментов может быть перенесена на молекулы АДФ.
Несмотря на большое число углеродных субстратов, доступных для сбраживания, количество реакций, приводящих непосредственно к синтезу АТФ при брожениях, сравнительно невелико. Наиболее распространены следующие из них:
1) ацетилфосфат + АДФ ® ацетат + АТФ;
2) 1,3-фосфоглицериновая кислота + АДФ ® 3-фосфоглицеринновая кислота + АТФ;
3) фосфоенолпировиноградная кислота + АДФ ® пировиноградная кислота + АТФ.
Другие реакции субстратного фосфорилирования ограничены какими-либо специфическими видами брожения. Например, сбраживание некоторых пиримидинов и аргинина, осуществляемое отдельными видами бактерий из рода Streptococcus, приводит к образованию карбамоилфосфата, фосфатная группа которого переносится на АДФ в реакции, катализируемой карбаматкиназой:
карбамоилфосфат + АДФ ® карбамат + АТФ.
Один вид клостридиев (C. cylindrosporum), сбраживающий пурины, способен образовывать формиат и тетрагидрофолат (ТГФК) из формилтетрагидрофолиевой кислоты в реакции, сопровождающейся фосфорилированием АДФ:
формил-ТГФК + АДФ + ФН ® формиат + ТГФК + АТФ.
Для этого вида указанная реакция служит основным путем получения АТФ.
Все реакции субстратного фосфорилирования локализованы в цитозоле клетки. Это указывает на простоту химических механизмов, лежащих в основе субстратного фосфорилирования.
Проблема акцептора электронов
Основная проблема всех процессов брожения — проблема акцептора электронов. В конечном итоге степень окисления и сопряженное с этим количество свободной энергии, а также характер образующихся продуктов определяются природой конечных акцепторов электронов. При брожениях конечными акцепторами электронов служат в основном органические соединения: метаболиты, образующиеся из исходных субстратов (пировиноградная кислота, ацетальдегид), или вещества, имеющиеся в среде культивирования (некоторые аминокислоты и другие органические соединения, способные восстанавливаться). В ряде брожений акцепторами электронов служат молекулы CO2, а также ионы водорода (H2). Кроме того, в отдельных случаях дополнительными акцепторами электронов могут быть некоторые достаточно окисленные неорганические соединения, такие как нитрат, молекулярная сера. Если конечным акцептором электронов является ацетальдегид, образуется этанол, если пируват — молочная кислота. Акцептирование электронов молекулами CO2 приводит у разных видов к возникновению формиата или ацетата, если же эту функцию выполняют ионы водорода, образуется молекулярный водород (H2).
Восстановленные соединения, акцептировавшие электроны, выделяются из клеток эубактерий в окружающую среду и накапливаются в ней в значительных количествах. Из-за низкого энергетического выхода процессов брожения для обеспечения энергией всех функций и биосинтетических процессов клетке приходится перерабатывать огромные количества субстратов.
Итак, брожение — это способ получения энергии, при котором АТФ образуется в процессе анаэробного окисления органических субстратов в реакциях субстратного фосфорилирования.
ГОМОФЕРМЕНТАТИВНОЕ МОЛОЧНОКИСЛОЕ БРОЖЕНИЕ
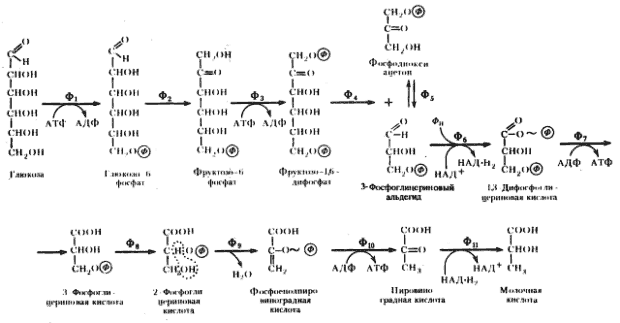
Последовательность биохимических реакций, лежащих в основе гомоферментативного молочнокислого брожения, получила название гликолитического пути (гликолиза)41, фруктозодифосфатного пути, или пути Эмбдена — Мейергофа — Парнаса (Н. Embden, О. Meyerhof, Я. О. Парнас), по именам исследователей, внесших больший вклад в изучение этого процесса. Общая схема гомоферментативного молочнокислого брожения представлена на рис. 53.
Рис. 53. Гомоферментативное молочнокислое брожение: Ф1 — гексокиназа; Ф2 — глюкозофосфатизомераза; Ф3 — фофсофруктокиназа; Ф4 — фруктозо-1,6-дифосфатальдолаза; Ф5 — триозофосфатизомераза; Ф6 — 3-ФГА-дегидрогеназа; Ф7 — фофсоглицерокиназа; Ф8 — фосфоглицеромутаза; Ф9 — енолаза; Ф10 — пируваткиназа; Ф11 — лактатдегиброгеназа (по Dagley, Nicholson, 1973) |