


Что такое молекулярный ламаркизм
Ю.В. Чайковский
Часто ламаркизмом называют признание наследования признаков, приобретенных организмом в течение жизни. Однако, как было отмечено в упомянутой статье Е.А. Ароновой, пресловутая «идея о наследовании приобретенных признаков стала известна как ламарковский механизм благодаря чему-то вроде исторической шутки». На самом деле в эту идею во времена Ламарка и Дарвина верили все. Суть же учения Ламарка состоит в другом – это «стремление природы к постепенному усложнению организации» и непрямое влияние внешних условий: «употребление того или иного органа или, наоборот, неупотребление его, вызывает изменение организации, которое передается потомству». Оба эти явления возможны в силу собственной активности особи, которая, тем самым, выступает у Ламарка как основной движущий фактор эволюции. Эти положения никогда не были опровергнуты опытом, они просто потеряли привлекательность в сравнении с более простой схемой неодарвинизма. «Идея о внутренней активности организма как факторе эволюционного развития все больше и больше игнорировалась», – пишет Аронова.
Попробуем еще раз сформулировать основные идеи ламаркизма как эволюционного учения.
1. Главным отличием, сделавшим ламаркизм популярным (особенно среди физиологов), явилось провозглашение в качестве ведущего фактора эволюции собственной активности особи.
2. Активность особи приводит к тому, что одни органы используются чаще и интенсивнее других, и это ведет их к преобразованию. Данный компонент теории Ламарка именуется «упражнение-неупражнение» (по-английски –«use-disuse»).
3. Наследование приобретенных признаков важно не само по себе, а как путь передачи итогов найденной удачной активности особи (например, упражнения органов) потомству.
4. Ламаркизм признает ведущим фактором эволюции также стремление организмов к совершенствованию (прогрессу). В этом утверждении проявляется наиболее резкое расхождение с дарвинизмом, полагающим прогресс всего лишь одной из форм приспособления к среде. Одновременно это утверждение близко идеям номогенеза – учения о закономерном характере путей эволюции.
5. С другой стороны, ламаркизм оказывается близок к дарвинизму и расходится с номогенезом в следующем. Номогенез видит своей целью выявление закономерностей эволюционных преобразований и утверждает, что они могут как быть, так и не быть связанными с функциями. Например, формы листьев вполне закономерны в том смысле, что выстраиваются в ряды2 , но во многих рядах не видно никакой связи с функциями: сходной формой листа могут обладать растения, весьма различные по экологическим особенностям и по происхождению, а у растений, сходных по экологии и/или состоящих в близком родстве, форма листьев может быть различной. Растущий в Москве клен и растущий на Кавказе платан имеют очень сходные контуры листьев, а в одной роще могут расти рядышком клен обыкновенный (платанолистный) и клен ясенелистный («американский клен»). Номогенез видит тут черту эволюции, тогда как ламаркизм и дарвинизм считают эволюционно значимыми лишь закономерности функционально значимых изменений.
6. Ламаркизм и дарвинизм рассматривают эволюцию как преобразование взрослых организмов в ходе развития жизни на Земле. Однако еще Этьен Жоффруа Сент-Илер, ученик Ламарка, развил концепцию, в которой ведущим фактором эволюции считается преобразование под действием изменений среды не взрослых особей, а зародышей. Этот взгляд позже был назван жоффруизмом.
До конца ХХ в. упоминания о новых взглядах на теорию ламаркизма в отечественной литературе были очень редки. Зато в последний год ушедшего века появились сразу две книги, осветившие, пусть и скупо, роль ламаркизма в новом эволюционизме.
Первая из них: Гродницкий Д.Л. Две теории биологической эволюции. (Красноярск, 2000). Две теории, о которых говорится в заглавии – это дарвинизм и эпигенетическая теория, так автор именует нынешний жоффруизм. Но и для ламаркизма, и, особенно, для номогенеза в книге тоже нашлось место. Автор говорит о двух положениях ламаркизма: наследовании приобретенных признаков и (правда, очень кратко) упражнении-неупражнении, но о прогрессе рассуждает только в контексте номогенеза.
Приведем цитату из книги.
«Общая схема опытов, подтверждающих возможность наследования приобретенных свойств, такова: особи или культуры клеток, обитающие в специфической среде, подвергаются кратковременному воздействию химического или физического фактора. После возвращения в прежние условия все особи и клетки (или большая их часть) обнаруживают новые признаки, которые передаются потомкам (Landman, 1991).
Один из примеров связан с блокированием активности генов путем их метилирования... Метиловый радикал, присоединенный к цитозину, изменяет конформацию молекулы ДНК, делая ген недоступным для регуляторных белков; при репликации ген копируется вместе с метиловой группой (Landman, 1991) ... появляются клетки других тканей, причем тканевая специфичность индуцированных клеток сохраняется на протяжении нескольких сотен поколений (Maynard Smith, Szathamary, 1995).»
(Видимо, речь идет о том, блокировка гена в клетке на начальной стадии индивидуального развития приводит к тому, что из такой клетки в дальнейшем формируется ткань с другими свойствами. – Прим. ред.)
Приведенный пример – простейшая иллюстрация молекулярного ламаркизма. Речь тут идет о наследовании длительном, но не навсегда, т.е. о длительных модификациях, известных генетикам уже 90 лет, но для которых лишь недавно был установлен молекулярный механизм. Длительные модификации, как эволюционный фактор, традиционно отвергаются дарвинизмом на том основании, что для эволюции якобы важны только генетические преобразования, носящие постоянный характер. Однако это соображение слабо, поскольку давно известно, что временные наследственные изменения могут заменяться на постоянные. В книге Гродницкого приведен длинный перечень таких изменений (у него они названы генетической ассимиляцией) и, в частности, результаты работ Г.Х. Шапошникова, который выращивал тлей на малопригодном для их питания субстрате (смертность – выше 99%) и примерно за 15 поколений (из которых почти все были партеногенетическими, получал различия ранга видовых3 . Правда, молекулярный механизм для объяснения этого конкретного явления еще не найден, имеются только гипотезы.
Читая Гродницкого, надо иметь в виду, что он полагает, будто при партеногенетическом размножении рекомбинация генетического материала отсутствует. На самом деле она идет (апомиксис), причем именно в этом случае возникает материал для сверхбыстрой эволюции.
Главное же, на мой взгляд, состоит в том, что новое появляется не за счет редких случайных мутаций, а в процессе массовой направленной изменчивости. На результат именно такой изменчивости и может действовать естественный отбор. К этому вопросу мы еще вернемся в п. 8 настоящей статьи.
Вторая из двух вышедших в России на рубеже веков книг, в которых затрагиваются вопросы ламаркизма: Голубовский М.Д. Век генетики: эволюция идей и понятий (СПб., 2000). К этой работе мы обратимся позже. Пока же отметим, что в ней можно найти описание механизма длительных модификаций и приведено еще несколько примеров молекулярного ламаркизма. Но, к сожалению, нигде прямо не сказано, что это ламаркизм.
2. Ламаркизм в нынешнем западном понимании
Основным полигоном для развития взглядов нового ламаркизма оказалась иммунология. Отметим однако, что за последние полвека сам смысл слова «иммунология» сильно изменился: прежние знания об иммунитете, об иммунитете врожденном, который общ всем организмам и наука о котором связана для нас с такими именами, как И.И. Мечников (животные) и Н.И. Вавилов (растения), теперь далеко отошли на периферию иммунологии. Разработанное 70–120 лет назад учение о врожденном иммунитете никем не оспаривается, но почти никем и не исследуется; всех занял анализ приобретенного иммунитета, связанного с изменением генов, происходящим у теплокровных животных и, прежде всего, млекопитающих, в течение жизни.
Главная тема книги трех австралийских иммуногенетиков во главе с Эдвардом Стилом «Что, если Ламарк прав?» (в русском переводе – М., 2002) – изложение взглядов авторов, видящих параллель между иммуногенезом, происходящим на протяжении жизни особи, и эволюцией, протекающей путем смены поколений, т.е. в историческом времени. Главное внимание уделяется синтезу гена, кодирующего иммуноглобулин – белок, выступающий в иммунной реакции как антитело, т.е как молекула, связывающая антиген – чужеродную молекулу. Синтез гена иммуноглобулина рассматривается в книге как модель синтеза всякого нового гена в эволюции.
Правда, на вопрос «Что такое ламаркизм?» и авторы, и редактор перевода ответили одинаково просто: наследование приобретенных признаков. Но это неверно. Как уже было сказано, тезис о наследовании приобретенных признаков – не положение какой-либо теории, а общее донаучное мнение, культурный феномен едва ли не у всех народов. Его не раз включали в свои построения многие мыслители, в том числе и до Ламарка, и после (например, Дарвин).
Данный тезис начал обсуждаться только в конце XIX в. в связи с работами Августа Вейсмана, заложившими основы неодарвинизма (в раннем смысле этого термина) – концепции, впервые провозгласившей невозможность наследования признаков, приобретенных особью в течение жизни, – поскольку, по Вейсману, наследственный материал половых клеток якобы не подвержен внешним воздействиям. Это «ненаследование» именуется в книге Э.Стила с соавторами «барьером Вейсмана».
Основная же суть ламаркизма никак не обсуждается. Упомянута, правда, тема «use-disuse», но весьма странно – как изобретение Ч.Дарвина (на самом деле он заимствовал ее у Ламарка). Вопрос о прогрессе даже не упоминается. Поэтому говорить, что авторы возрождают ламаркистские взгляды, можно лишь с радикальной оговоркой: в первую очередь в книге реанимируется ламаркистский элемент учения Дарвина.
Надо заметить, что подобное (непозволительное!) сужение понимания ламаркизма – тоже культурно-исторический феномен. В самом деле, авторы приводят цитаты из пяти нынешних англоязычных биологических словарей, и во всех ламаркизм подан именно как наследование приобретенных признаков. Разумеется, и по-английски можно прочесть о ламаркизме кое-что более содержательное, но такое узкое понимание этой теории на самом деле преобладает.
Кстати, вся обширная библиография книги англоязычна. Англоязычная же литература, в достаточной степени охватывающая работы в области молекулярной биологии, далеко не так полна в отношении эволюционных идей вообще и ламаркизма в особенности. Тут важнее работы, вышедшие на французском, немецком и русском языках. Поэтому неудивительно, что заглавие книги плохо соответствует ее содержанию, что анализа проблем эволюции в ней фактически нет.
Наши популярные издания были и остаются лучше. Вот, к примеру, статья в обычном энциклопедическом двухтомнике для широкого читателя (Российский энциклопедический словарь. – М., 2000).
«Ламаркизм, первая целостная концепция эволюции живой природы, сформулированная Ж.Б. Ламарком. По Ламарку, виды животных и растений постоянно изменяются, усложняясь в своей организации в результате влияния внешней среды на организм некоего внутреннего стремления всех организмов к усовершенствованию. В дальнейшем ламаркизм подвергался резкой критике сторонниками дарвинизма, но вместе с тем находил поддержку в различных направлениях неоламаркизма».
Как видим, наследование приобретенных признаков даже не упомянуто. Его находим в другой статье двухтомника.
«Неоламаркизм, совокупность разнородных концепций в эволюционном учении, возникших во 2-й половине XIX в. в связи с развитием отдельных положений ламаркизма. Механоламаркизм приписывал ведущую роль в эволюции условиям внешней среды; ортоламаркизм усматривал основную причину развития во внутренних свойствах организмов, предопределяющих прямолинейный характер эволюции; психоламаркизм считал основным источником эволюции сознательные волевые акты организмов. Общее для всех этих концепций – признание наследования приобретенных признаков и отрицание формообразующей роли естественного отбора».
Но вернемся к книге Э.Стила с соавторами. Итак, замахнувшись на проблему в целом, австралийские иммунологи на самом деле рассуждают только об одной из черт неоламаркизма. Но при этом они приводят много интересных фактов. Рассмотрим их, а затем сами подумаем, что нового можно извлечь из этого материала для понимания эволюции.
3. Ламаркизм и дарвинизм
Первая глава книги названа: «Идеи Ламарка и Дарвина – две стороны одной медали». В ее начале авторы справедливо отмечают: «Революционные представления Чарлза Дарвина о естественном отборе как главной движущей силе эволюции сейчас превратились в догму. Обновление наших взглядов на эволюцию требует учета данных, полученных молекулярной генетикой, особенно – молекулярной генетикой иммунной системы». Это верно: все известные мне руководства по дарвинизму оперируют понятиями пятидесятилетней давности; а если там время от времени и используются новые термины, то не по существу.
Далее авторы напоминают: «В научной картине «внутреннего мира» клеток и молекул иммунной системы неоламаркистские представления об обратной связи генов сомы и зародышевой линии (Т.е. геномов клеток тела – соматических и геномов половых клеток. – Прим. ред.) давно занимают видное и законное место»». Тут они правы: с 1970-х гг. растет уверенность в недостаточности концепции случайных мутаций для понимания хода внутриклеточного наследственного приспособления. В частности, открытие у бактерий способности включать в свой геном дополнительный генетический материал привело к пониманию того, что случайные ненаправленные мутации вносят лишь малый вклад в явление лекарственной устойчивости – устойчивые к лекарствам штаммы бактерий возникают слишком часто и при этом сразу, а не постепенно.
И хотя именно на бактериях был в 1943 г. получен главный для неодарвинизма результат: устойчивость к антибиотикам появляется только за счет спонтанных мутаций; на тех же бактериях американский генетик Джон Кэйрнс (Cairns) в 1988 г. этот результат опроверг, показав, что среди мутаций присутствуют индуцированные, т.е. вызванные условиями опыта. Этот хорошо известный факт описан, в частности, в упомянутой выше книге М.Голубовского. Писал об этом и я (Эволюция. Часть 5)4 .
Оказалось, что в опытах 1943 г. выживали те бактерии, которые уже до начала эксперимента имели ген, обеспечивающий устойчивость к антибиотику. А в опытах Кэйрнса, кроме уже существовавших мутантов, нашлись и такие, которые возникли именно в ответ на действие нового внешнего фактора. Мутации оказались случайными, но направленными, и Кэйрнс смог лишь воскликнуть: «Поразительно, сколь малообоснованным было общепринятое мнение».
Если внимательно проанализировать первоначальные опыты, то окажется, что сама их постановка не давала возможности получить что-либо, кроме «доказательства» случайности мутаций. В частности, в них всегда использовались столь высокие концентрации ядов, что возможность физиологического приспособления бактерий практически исключалась. Хотя вроде бы ставилась цель – проверить, идет ли таковое приспособление и носит ли оно наследственный характер. Когда же смертельные дозы были уменьшены до стрессовых, тут же обнаружились акты наследуемого приспособления.
Вскоре такие же результаты были получены и на более сложных организмах, причем всюду дело было действительно в физиологии, в стрессе. По этому поводу Голубовский в своей книге пишет: «Как будто клетки в условиях жесткого стресса, не делясь (!), вели генетический поиск и адаптивно меняли свой геном».
О генетическом поиске мы поговорим позже – в п. 8 (и узнаем, что он в самом деле способен идти в неделящейся клетке), а сейчас обращу ваше внимание на стресс: одним из самых важных достижений эволюционизма последнего полувека стало понимание того, что клетка в состоянии стресса способна изменять то, что в обычных условиях неизменно, – свою генетическую информацию. Или, в терминах Стила с соавторами, преодолевать барьер Вейсмана.
Тут я должен предупредить читателей: при чтении книги австралийских иммунологов остается непонятным, зачем вообще опыты Кэйрнса были нужны, и в чем состоит спор, поскольку авторы уверяют, что у одноклеточных барьера Вейсмана нет. И огорошивают читателя: «А что же с растениями? У них нет барьера Вейсмана, отделяющего сому от зародышевой линии. Приобретенные соматические модификации растений, связанные с изменениями генов, могут, в принципе передаваться потомству ... Итак, это уже не секрет: эволюция по Ламарку была и есть, это факт из жизни растений!»
Позвольте, если дело обстоит так, то растениям надо бы посвятить основную часть книги, а не этот единственный абзац. Если бы «барьер Вейсмана» существовал лишь у животных, то весь неодарвинизм был бы учением об особенностях эволюции животных. На самом деле авторы спутали учение Вейсмана о зародышевой плазме с теорией зародышевого пути. Такового пути у растений и губок действительно нет5. К сожалению, подобных несуразностей в книге довольно много.
Главную же мысль первой главы книги «Что, если Ламарк прав?» можно выразить так: дарвинизм признает только случайную, ни от чего не зависящую изменчивость, а ламаркизм утверждает, что «генетическая изменчивость возникает одновременно с отбором». В этом авторы правы. Однако, по их мнению, данный феномен «легко объясняет, почему некоторые виды смогли очень быстро генетически измениться при внезапных изменениях среды, во время катастроф; также легко объяснить быстрое создание разных пород домашних животных». Нет, до объяснения тут еще далеко: не указан способ (механизм) приобретения наследственных изменений. Однако и о пути их наследования вполне стоит поговорить.
4. Вейсман против Вейсмана
Впервые Э.Стил выступил со своей эволюционной концепцией еще в 1979 г. (см. статью Е.Ароновой) и начал тогда с анализа «центральной догмы молекулярной биологии». Это утверждение первоначально (1958 г.) записывалось так:
ДНК - -> ДНК - -> РНК - -> белок
и гласило, что белок синтезируется только на РНК-вой матрице, РНК – только на ДНК-вой, а ДНК реплицирует саму себя. Однако вскоре (1970 г.) оказалось, что на РНК-вой матрице может синтезироваться ДНК – это явление называется обратной транскрипцией. Кроме того (это было ясно давно), синтез нуклеиновых кислот требует, кроме полинуклеотидной матрицы, еще и участия белков. Пусть матрицей белок и не служит, но изменение белковых текстов способно повлечь изменение текстов и ДНК, и РНК, и самих белков.
Вдобавок, в 1982 г., Ф.Альт и Д.Балтимор открыли нематричный синтез ДНК: в ходе синтеза гена, кодирующего антитело, идет сшивка фрагментов прежних генов, причем в точке сшивки в текст ДНК встраивается небольшой (кодирующий до восьми аминокислот) фрагмент, ни в какой матрице не хранимый, а синтезируемый и встраиваемый ферментативно. Насколько сейчас известно, для работы антитела информативна только длина этой вставки (а не состав ее!), но нам важнее то, что данная генетическая информация взята не из генетического текста, а из функционального состояния ферментативной системы. В этом смысле она перенесена с белков на ДНК и означает переход: белок - -> ДНК.
Затем появились и другие примеры нарушения «центральной догмы», и в возникшей по этому поводу дискуссии6 мы видим традиционные взаимные упреки в неверном понимании самой догмы. Так, ее сторонник сетует на приемы «бесчестной компрометации», когда «тезису, который надо скомпрометировать, преднамеренно приписывают формально схожее, но на самом деле умышленно ложное и весьма часто расширительное утверждение». А противник догмы утверждает прямо противоположное: «Под давлением фактов начался отход от позиции «барьера Вейсмана». Однако прямо признаться в этом сторонники гипотезы Вейсмана не желали и стали менять формулировки, лишь бы сохранить на словах саму эту гипотезу».
Обвиняя друг друга в сознательном обмане, добиться успеха невозможно. Поскольку всякий ученый считает себя правым вполне искренне, то надо стараться друг друга понять. Да, подмена понятий при обсуждении центральной догмы имеет место, но со стороны как ее защитников, так и противников. Обе стороны признают, что догма верна в ее самом узком смысле – в смысле отсутствия «обратной трансляции»: белок не служит матрицей для нуклеиновой кислоты. Далее обе же стороны согласны в отрицании догмы в ее самом широком смысле – как запрет влияния белков на синтез нуклеиновых кислот. Однако если сторонники догмы делают из этих двух утверждений вывод, что догма истинна, то противники – что она ложна. Учтя сказанное, мы примем такую позицию: догма в самом узком смысле верна, но неинтересна, поэтому надо выяснить, в каком смысле, в какой мере и какими конкретными механизмами осуществляется влияние белков (точнее, всей физиологии клетки) на генетические тексты. Для понимания эволюции нужно это, а не взаимные упреки.
Обсуждению нынешнего состояния центральной догмы посвящена, в частности, упомянутая подборка статей в журнале «Химия и жизнь» за 2003 г. Четыре из пяти опубликованных работ убеждают читателя, что от центральной догмы мало что осталось, а автор еще одной уверен, что ставить опыты нет нужды, ибо приобретенные признаки не могут наследоваться в принципе.
С того утверждения, что центральная догма являет собой молекулярную формулировку запрета наследования приобретенных признаков, и начал в 1979 г. свою работу Э.Стил. Его исходная мысль была парадоксальна: хотя «центральная догма» ничем не доказана и препятствует пониманию эволюции, но путь к новой теории лежит именно через уяснение сути этой догмы. А поскольку она – молекулярный аналог идеи зародышевой плазмы Вейсмана, то начать надо с анализа его учения. «Моя точка зрения такова: основной блок любого понимания наследования приобретенных признаков можно найти прямо в доктрине Вейсмана», – заявлял тогда Стил.
В самом деле, Вейсман сам был вынужден отказаться от крайних своих утверждений. В 1892 г. он писал: «Корень наследственных изменений должен лежать глубже (чем в слиянии родительских половых клеток. – Ю.Ч.) и заключаться в прямом воздействии внешних влияний на биофоры (единицы наследственности – Ю.Ч.)». В чем же состоит такое воздействие? Этого, разумеется, Вейсман не знал и, в стиле натурфилософов прошлого, написал: «Начало изменения основывается на малых неправильностях в питании зародышевой плазмы». Тут-то Стил и смог дать радикальное уточнение, невозможное в дни Вейсмана: «питание» наследственного материала состоит в усвоении им генетического материала некоторых вирусов.
Конечно, сводить эволюцию, даже только приспособительную, к одному лишь наследованию приобретенных признаков наивно, зато такое сужение задачи позволило Стилу сразу вчерне решить ее. По его мнению, зародышевая плазма действительно играет ведущую роль в эволюции, поскольку ее фрагменты передаются между органами и организмами с помощью вирусов. Тем самым, вирусы, по крайней мере некоторые их формы, например ретровирусы (вирусы, РНК которых передает свою информацию в хромосому организма путем обратной транскрипции), являются не только и даже не столько причиной болезней, сколько необходимым эволюционным агентом.
Главная мысль новой книги Стила с соавторами проста и убедительна: синтез антител есть создание новой генетической информации, не сводимое к случайным ненаправленным вариациям «по Дарвину», причем нужный ген формируется целенаправленно. Тем самым феномен приобретенного пожизненного иммунитета выступает как несомненный факт наследования приобретенного признака, причем наследование происходит путем включения вновь найденного гена, кодирующего антитело, в хромосомы клеток иммунной памяти, из которых при надобности вновь производятся клетки, способные производить антитела.
Исходя из этого факта делается правдоподобное допущение: эволюционно значимая информация может передаваться из хромосом стволовых клеток лимфоидной системы в хромосомы стволовых клеток половой железы. А осуществляют эту передачу, по Стилу, ретровирусы. Это допущение еще предстоит проверить. Пока же поговорим о механизме приобретения новой генетической информации в ходе иммуногенеза и подумаем, что это дает для понимания эволюции.
5. Генетический принцип обеспечения разнообразия антител
У зародыша млекопитающих есть совсем немного генов, кодирующих иммуноглобулины, – около сотни. Их последующее разнообразие в ходе развития организма каждый раз создается заново, точно так же, как заново создается любой орган. Создается оно путем комбинирования фрагментов существующих генов. Но этого разнообразия оказывается мало, поэтому конкретное антитело обычно не выбирается из наличных, а создается в ответ на конкретную заразу (на антиген). Процесс идет так.
В стрессовой ситуации, которую создает вторжение антигена, включается механизм перестройки генов иммуноглобулинов: генетическая система по каким-то не вполне еще понятным правилам режет и сшивает фрагменты генов до тех пор, пока не найдет приемлемый вариант – тот, что синтезирует антитело, которое реагирует с вторгшимся антигеном. Найденный вариант клонируется (т.е. размножается из единственного родоначального экземпляра).
За открытие этого механизма иммунолог из Японии Сусуму Тонегава получил в 1987 г. Нобелевскую премию (работа начата в Швейцарии, а завершена в США). Суть открытия в том, что ген может быть переделан в цитоплазме.
Указанный механизм рекомбинаций поставляет антитела, связывающие антигены довольно слабо. Для улучшения их «качества», для тонкой подстройки, осуществляется следующий этап, соматический (т.е. не связанный с размножением), – гипермутагенез. Включаясь после создания нужной комбинации фрагментов он осуществляет превращение «черновой болванки» в «готовое изделие».
Гипермутагенез заключается в том, что при клонировании гены «болванки» (первично найденного варианта) мутируют с огромной частотой (каждый тысячный нуклеотид заменяется, тогда как обычно точковый мутагенез в 100 миллионов раз менее интенсивен), а потом с их копий синтезируется масса чуть отличных друг от друга белковых цепей (рис. 1) антител, какое-то из которых оказывается подогнанным к антигену наилучшим образом. Этот окончательный вариант снова клонируется и запоминается клетками иммунной памяти, т.е. наследуется на время жизни особи (возникает приобретенный иммунитет).
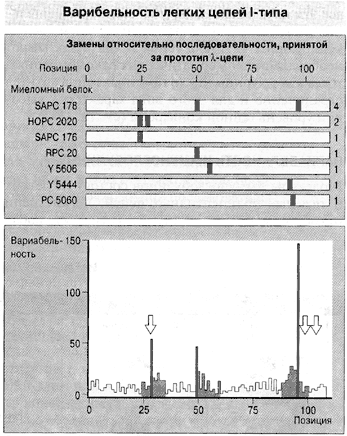
Рис. 1. Избирательность гипермутагенеза (эффект Ву–Кэбота), которую установили в 1970 г. Тай Тэ Ву и Элвин Кэбот на легкой цепи одного из иммуноглобулинов мыши. (По Ройт и др., 2000.) Сходная картина получена и на других объектах. Вверху: 7 фрагментов белка с удачными заменами аминокислот. (Цифры справа – число необходимых для этого замен нуклеотидов.) Внизу: частота наблюдавшихся в опыте замен аминокислот
В этом, грубо говоря, состоит генетический принцип обеспечения разнообразия антител (термин Тонегавы): возникшие при перестройках фрагменты сшиваются (механизм Тонегавы), причем с нематричными вставками (механизм Альта–Балтимора, п.4), затем успешный вариант точно подгоняется к антигену (механизм гипермутагенеза), клонируется и запоминается (соматическое наследование).
Словом, гены антител образуются не за счет случайных мутаций, как думали прежде, а путем многостадийного процесса (рис. 2), в котором лишь одну ступень можно назвать мутагенезом и то в особом смысле: он направлен – в том смысле, что происходит только в нужных участках нужных генов, зато с неимоверной частотой.
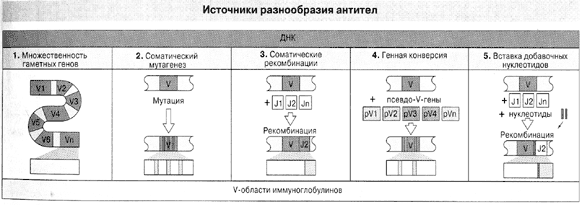
Рис. 2. Пять источников разнообразия антител ( По Ройт и др., 2000).
Кроме описанных в тексте, здесь указана генная конверсия, смысл которой тот же, что у рекомбинации по Тонегаве (комбинируются гипервариабельные участки V), но добавочными элементами комбинирования служат не соединительные сегменты J, а псевдогены, т.е. гены, утратившие собственную активность
Сам Тонегава в нобелевской лекции ясно провел параллель иммуногенеза с эволюцией: «Подобно организмам в экосистеме, эти лимфоциты – субъекты отбора антигенами, и приспособленнейший будет выживать. И... иммунную систему индивида можно рассматривать как своего рода Дарвинов микрокосм».
Параллель очевидна, но надо выяснить, причем тут Дарвин. Оказывается, иммуногенез схож не с естественным отбором, а с искусственным. Мы подошли к самому важному пункту.
6. Роль отбора в иммуногенезе
Мир учения Дарвина – это мир малых вариаций, последовательно вытесняющих друг друга в борьбе за дефицитные ресурсы. Только при таком понимании естественного отбора Дарвину удалось построить свою теорию. И хотя позже было предложено множество иных пониманий, годились они только для своих узких конкретных целей. Поэтому важно понять, насколько справедлива аналогия Тонегавы (не раз повторенная другими).
Легко видеть, что в иммунногенезе все совсем не так, как у Дарвина: клетка делится не потому, что получила какой-то жизненный ресурс или пришло время, а по «приказу» (сходному с «приказом» селекционера); и этот «приказ» к размножению дает клетке ее антитело, связавшееся с антигеном. Далее, малые вариации являют здесь только одну из ступеней изменчивости, тонкую подстройку.
Как только в крови появились «болванки», связанные с антигеном, тут же возникают (в лимфатических узлах и в селезенке) центры размножения, где «болванки» накапливаются, и тем самым накапливается антиген, который связан, но не разрушается. Только здесь и идет гипермутагенез («Что, если Ламарк прав?», с. 133).
Это очень похоже на ферму. Сравнения клеток по выживаемости тут нет: словно селекционер на ферме, иммунная система колоссально размножает тех, кто несет желаемый признак, и вовсе не допускает к размножению остальных. Существенно, что эти остальные тут же уничтожаются (механизмом апоптоза, т.е. самоуничтожения клетки), чего в дарвиновском естественном отборе тоже нет.
Что касается конкуренции за ресурсы, то вопрос сложнее: в одном смысле иммунологи ее допускают – в процедуре точной подстройки. Эту процедуру они называют повышением аффинности В-клеток, т.е. клеток, производящих антитела. (Слово «аффинность» означает по-русски «сродство»; им обозначают специфичность и силу связывания антитела с антигеном.) Авторы нового английского руководства: Ройт А. и др. Иммунология (М., 2000) в параграфе «Созревание аффинности зависит от селекции клеток» указывают: «Степень повышения аффинности обратно пропорциональна дозе введенного антигена: при введении высоких доз сродство возрастает слабее, чем после иммунизации низкими дозами антигена. Высказано предположение, что при низкой концентрации антигена его связывают в достаточном количестве лишь те В-клетки, которые обладают высокоаффинными рецепторами... При значительной же концентрации антигена его связывают и активируются как высоко-, так и низкоаффинные В-клетки».
Предположение вполне естественно, но это пока всего лишь предположение, и спасибо авторам, честно отметившим этот факт (другие, в том числе Стил, пишут иное – что В-клетки в самом деле конкурируют за дефицитный антиген, так что между ними якобы идет настоящий естественный отбор по Дарвину). Для выявления конкуренции нужны специальные опыты. В частности, необходимо определить те концентрации антигена, при которых эффект малых доз возникает и при которых исчезает, а затем выяснить, имеет ли место в этих пределах дефицит антигена.
Пока это не сделано, налицо лишь сам эффект малых доз, а он известен всем врачам и физиологам. Говорят даже, что всякое лекарство есть яд, даваемый в малых (стимулирующих) дозах. Это значит, что с ростом дозы меняется не только сила эффекта, но и сам эффект может смениться на противоположный, а это (смену знака процесса) вряд ли возможно связать, не напрягая фантазии, с какой-либо конкуренцией.
Словом, «Дарвинов микрокосм» сомнителен, и Тонегаве лучше было бы сказать: «микрокосм Эмпедокла» – об этом античном мыслителе (полагавшем, что первые организмы произошли путем случайной комбинации органов, но потомство оставили только те, кто был способен жить) можно прочесть в любом курсе истории эволюционных учений. Иммуногенез на первом этапе синтеза гена антитела осуществляется именно путем комбинирования блоков. И, поскольку иммунитет на самом деле работает (а эволюционные доктрины только строят воображаемые картины), постольку знание о нем как раз и может служить для оценки справедливости эволюционных доктрин и для их коррекции.
7. Иммунология и общие проблемы эволюции
Заглянем в Биологический энциклопедический словарь.
«Иммунология – биологическая наука, изучающая защитные реакции организма, направленные на сохранение его структурной и функциональной целостности и биологической индивидуальности».
Напомню: именно так, широко, предложил в 1964 г. понимать иммунологию Фрэнк Макфарлейн Бернет, основатель австралийской иммунологической школы и нобелевский лауреат. С этой позиции борьба с заразой – лишь одна из задач иммунитета. Другая задача – контроль совместимости тканей организма.
Как уже сказано выше, иммунитет есть у всех организмов, и они все жизнеспособны, следовательно простого (врожденного) иммунитета для успешной борьбы с инфекцией достаточно. Но на это обращают внимание лишь немногие (например: С.Н. Румянцев. Микробы, эволюция, иммунитет. – Л., 1984), а в книге Э.Стила с соавторами об этом нет ни слова.
Хотя А.Ройт с соавторами в своем цитированном выше учебнике («Иммунология») врожденный иммунитет едва упоминают, но в эволюционной главе заявляют прямо: «Беспозвоночные лишены лимфоцитов и антител, но тем не менее обладают эффективными механизмами защиты. ... Внедрение патогенных организмов и паразитов вызывает у беспозвоночных быстрый иммунный ответ, что и обеспечивает громадное разнообразие и изобилие этих животных». И ставят вопрос: «только позвоночные обладают лимфоцитами с высокой специфичностью и среди них клетками иммунологической памяти. Какие же факторы внешней среды могли обусловить усложнение иммунной системы у позвоночных?»
Вот первый вопрос, связывающий иммунологию с эволюцией. В учебнике ответа не дано, и у меня его тоже нет, но замечу, что факторы вовсе не обязательно искать только во внешней среде – не менее важна внутренняя среда организма.
Далее, у беспозвоночных есть крупные формы, например головоногие. Они не могут (как могут мелкие организмы) компенсировать высокую смертность от болезней высокой рождаемостью, а потому нуждаются в сохранении особей не меньше, чем мы с вами, и гораздо больше, чем мыши и те мелкие звери, из которых мы произошли. Более того, самые крупные и медленно размножающиеся на Земле организмы – гигантские деревья, а они обладают самым простым типом иммунитета. (Следует помнить, что возможность компенсации высокой смертности высокой рождаемостью – число производимых особью потомков, их выживаемость и скорость полового созревания – далеко не всегда связана напрямую с таким неспецифическим признаком, как размер организма. – Прим. ред.)
Добавим, что недавние исследования показывают, что у растений тоже наблюдается высокая генетическая вариабельность элементов иммунной системы, сходная с вариабельностью генов наших антител7 . Но растения не страдают от наших автоиммунных болезней (рак, рассеянный склероз, аллергия и т.п.), при которых иммунная система буквально поедает хозяина.
Этим ставится второй вопрос: зачем нам наш устрашающе сложный иммунитет, ежечасно готовый превратиться из защитника