


Фотосинтез как основа энергетики биосферы
Содержание
1 Фотосинтез. Определение, общее уравнение, основные этапы становления учения о фотосинтезе. Историческое значение работ К.А. Тимирязева
2 Фотосинтез как основа энергетики биосферы. Космическая роль фотосинтеза. Роль фотосинтеза в процессах энергетического и пластического обмена растительного организма
3 Структурная организация фотосинтетического аппарата
3.1 Лист как орган фотосинтеза
3.2 Состав, строение, функции, возникновение и развитие хлоропластов
4 Пигменты хлоропластов
4.1 Хлорофиллы: состав, структура, биосинтез, оптические и химические свойства, значение
4.2 Каратиноиды. Их структура функции и физиологическая роль
4.3 Фикобилины. Их структура функции и физиологическая роль
4.4 Экологическое значение спектрально-различных форм пигментов у фотосинтезирующих организмов
5 Световая фаза фотосинтеза
5.1 Фотофизический этап. Электронно-возбуждённое состояние пигментов. Представление о фотосинтетической единице. Антенные комплексы. Реакционные центры. Преобразование энергии в реакционном центре
5.2 Фотохимический этап Электронно-транспортная цепь фотосинтеза. Представления о функционировании двух фотосистем. Фотофосфорилирование. Системы фотоокисления воды и выделения кислорода при фотосинтезе. Фотофосфорилирование. Связь фотосинтетической ассимиляции СО2 с фотохимическими реакциями
6 Метаболизм углерода при фотосинтезе (темновая фаза)
6.1 Химизм реакции цикла Кальвина
6.2 Цикл Хэтча-Слэка-Карпилова, его эволюционное значение. Различные типы усвоения углекислого газа С4-растениями
6.3 САМ-тип метаболизма. Потоки метаболитов в хлоропласт и из него
7 Фотодыхание, его значение. Сравнение фотодыхания у растений с различными типами метаболизма углерода
8 Эндогенные механизмы регуляции процесса фотосинтеза
9 Зависимость процесса фотосинтеза от факторов внешней среды
Список литературы
ВВЕДЕНИЕ
Фотосинтез — это процесс трансформации поглощенной организмом энергии света в химическую энергию органических (и неорганических) соединений. Главную роль в этом процессе играет использование энергии света для восстановления С02 до уровня углеводов. Часто говорят о фототрофной функции фотосинтеза, понимая под этим использование энергии света в различных эндергонических реакциях в живом организме. Фотосинтез осуществляют высшие растения, водоросли и некоторые бактерии. Он играет определяющую роль в энергетике биосферы.
1 Фотосинтез. Определение, общее уравнение, основные этапы становления учения о фотосинтезе. Историческое значение работ К. А. Тимирязева
С древних времен люди отмечали, что деревья могут вырастать на бесплодных скалах. Английский ботаник и химик С.Гейлс в своей книге «Статика растений» (1727) высказал предположение, что растения значительную часть «пищи» получают из воздуха. При этом он вслед за И.Ньютоном полагал, что свет, поглощаемый листьями, «облагораживает» эту «пищу». Сходные мысли находят у М.В.Ломоносова. В «Слове о явлениях воздушных» (1753) он писал: «Преизобильное ращение тучных дерев, которые на бесплодном песку корень свой утверждали, ясно изъявляет, что листами жирный тук из воздуха впитывают...». Так возникла идея о воздушном питании растений.
Началом экспериментальных работ в области фотосинтеза послужили опыты английского химика Дж.Пристли, который в 1771г. обнаружил, что растения мяты, помещенные в стеклянный кувшин, опрокинутый в сосуд с водой, «исправляют» в нем воздух, «испорченный» горением свечи или дыханием мыши. Свеча могла длительно гореть, а мышь дышать, если под стеклянным колпаком находились зеленые растения. В связи с этими и последующими опытами Пристли в 1774 г. открыл кислород. Через год независимо от него кислород был открыт во Франции А.Л.Лавуазье, который и дал название этому газу.
В 1776 г. шведский химик К.В.Шееле повторил опыты Пристли, но не получил того же эффекта, который был описан Пристли. Растения в опытах Шееле делали воздух непригодным для дыхания так же, как горящая свеча. Объяснил противоречие между результатами опытов Пристли и Шееле голландский врач Я.Л.Ингенгауз. Он обнаружил, что все зависит от света: Пристли ставил опыты при ярком свете, Шееле — в помещении с недостаточным освещением. Ингенхауз показал, что зеленые растения выделяют кислород только при действии на них света. Зеленые растения в темноте, а их незеленые органы (например, корни) в темноте и при освещении поглощают кислород точно так же, как животные в процессе дыхания.
Швейцарский естествоиспытатель Ж.Сенебье в 1782 г. установил, что растения на свету не только выделяют кислород, но и поглощают испорченный воздух, т. е. С02. Сенебье назвал поглощение С02 «углеродным питанием».Применив методы количественного анализа, швейцарский ученый Т.Соссюр в 1804 г. показал, что растения на свету действительно усваивают углерод С02, выделяя при этом эквивалентное количество кислорода. Однако нарастание сухой массы растений превышает прирост количества углерода. Это превышение было значительно большим, чем количество поглощенных минеральных веществ. Соссюр сделал вывод, что органическая масса растения образуется не только за счет С02, но и за счет воды, т. е. вода такой же необходимый элемент питания, как и диоксид углерода. Результаты этих опытов были тщательно проверены французским агрохимиком Ж.Б.Буссенго (1840), который полностью подтвердил данные Соссюра.
Французские химики П.Ж..Пельтье и Ж..Каванту в 1817 г. выделили из листьев зеленый пигмент и назвали его хлорофиллом (от греч. «chloros» — зеленый и «phyllon» — лист), который, как выяснилось позднее, целиком локализован в хлоропластах. В 1865 г. немецкий физиолог растений Ю.Сакс продемонстрировал, что на свету в листьях образуется крахмал и что он находится в хлоропластах. Опыты ставили следующим образом. Листья предварительно выдерживали в темноте, затем освещали половинку каждого листа, а другую половинку, закрытую плотным картоном, оставляли в темноте. После экспозиции листья обесцвечивали спиртом и обрабатывали раствором йода. Освещенные части листьев становились темно-фиолетовыми из-за образования комплекса крахмала с йодом, а затемненные участки оставались неокрашенными. Микроскопический анализ показал, что крахмальные зерна образуются именно в хлоропластах. Эта «проба Сакса», как ее стали называть, настолько чувствительна, что на листьях удается получить отпечатки с фотонегативов. На прямом солнечном свету для образования крахмала достаточно уже 5 мин. А.С.Фаминцын в 60-е годы прошлого столетия наблюдал образование крахмала в клетках водоросли спирогиры уже через 30 мин освещения слабым светом керосиновой лампы.
Первые опыты по выяснению места образования кислорода при фотосинтезе были сделаны немецким физиологом Т.В.Энгельманом (1881). Нить спирогиры помещали в камеру с водой или в висячую каплю. Края покровного стекла, которым закрывали ячейку, замазывали вазелином, чтобы кислород воздуха не проникал из внешней среды. В эту же камеру помещались бактерии, способные двигаться только в присутствии кислорода. В темноте водоросль дышала и кислород исчезал из камеры. Затем включается свет и в ходе фотосинтеза выделяется кислород. Оказалось, что при включении света все бактерии «оживали» и скапливались вокруг локально освещенных участков хлоропластов. Следовательно, именно хлоропласты выделяют кислород на свету.
Вопросом о роли света в процессах фотосинтеза начали заниматься с середины XIX в. Американский физик Дж.У.Дрепер в 1846 г , вслед за ним Ю.Сакс и В.Пфеффер считали, что фотосинтез лучше всего осуществляется в желтых лучах, наиболее ярких для человеческого глаза. В то же время было уже хорошо известно, что хлорофилл имеет резко выраженные максимумы поглощения в красной и синей частях спектра. Исходя из этих данных делался вывод, что процесс фотосинтеза не подчиняется закону сохранения энергии, а свет действует на зеленый лист как раздражитель. Против этого утверждения выступили Ю.Р.Майер и Г.Гельмгольц, сформулировавшие закон сохранения и превращения энергии. На основе чисто теоретических рассуждений они утверждали, что зеленые растения поглощают лучистую солнечную энергию и превращают ее в химическую, т. е. фотосинтез — это процесс трансформации энергии света в энергию химических связей. Однако экспериментальных доказательств этой точки зрения не было. Они были получены крупнейшим физиологом растений К.А.Тимирязевым, который изложил их в своей докторской диссертации «Об усвоении света растением» (1875). Тимирязев нашел, что слишком широкие щели при пропускании света, разложенного с помощью призмы, не обеспечивают его монохроматической чистоты. Этот недостаток в постановке опытов и привел Дрепера и Пфеффера к неверным выводам. Тимирязев использовал в работе очень узкие щели и помещал в полоски монохроматического света тонкие пробирки с высечками из листьев. Для этих опытов ему пришлось разработать очень чувствительные методы газового микроанализа. В результате было установлено, что интенсивность ассимиляции С02 максимальна при освещении листьев красным светом, т. е. тем светом, который в наибольшей степени поглощается хлорофиллом.
Тимирязев постулировал, что при ассимиляции С02 хлорофилл служит оптическим сенсибилизатором (т. е. веществом, увеличивающим чувствительность к свету) и что он непосредственно участвует в процессе фотосинтеза, необратимо переходя из восстановленного состояния в окисленное. Он сформулировал также идею о космической роли фотосинтеза: фотосинтез — единственный процесс, с помощью которого космическая солнечная энергия улавливается и остается на Земле, трансформируясь в другие формы энергии. Тимирязев писал, что в хлоропласте лучистая энергия солнечного света превращается в химическую энергию углеводов. Крахмал, клейковина и другие соединения, консервирующие солнечную энергию, служат нам пищей. Освобождаясь в нашем теле в процессе дыхания, эта энергия солнечного луча согревает нас, приводит в движение, поддерживает мышление.
Результаты изучения воздушного питания растений за первые сто лет после опытов Пристли нашли свое выражение в общем уравнении фотосинтеза:
свет
6СО2 + 12Н2О --- С6Н12О6 + 6СО2 + 6Н2О
хлорофилл
Происхождение 02 при фотосинтезе. В приведенном выше уравнении остается неясным, каково происхождение выделяющегося кислорода (из С02 или из Н20). Во второй половине XIX в. и в начале XX в. считалось, что при фотосинтезе происходит фоторазложение С02 с последующим восстановлением углерода до углеводов с участием воды. Немецкий химик А.Байер в 1870 г. предложил следующий порядок реакций:
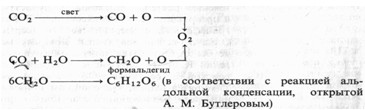
Исходя из этой гипотезы разложения диоксида углерода, можно было предположить, что для образования сахаров и крахмала растения должны усваивать промежуточные продукты — СО или формальдегид. Однако оба соединения оказались токсичными для растений, что указывало на ошибочность схемы Байера.
В 1893 г. крупнейший русский биохимик А.Н.Бах высказал мысль о том, что ассимиляция С02 при фотосинтезе связана не с отщеплением 02 от диоксида углерода, а является сопряженным окислительно-восстановительным процессом, происходящим за счет водорода и гидроксила воды, причем 02 выделяется из воды через промежуточные перекисные соединения.
![]() Голландский микробиолог К.Б. ван Ниль, изучая особенности бактериального фотосинтеза и сравнивая его с фотосинтезом у растений, в 1937—1941 гг. также пришел к заключению, что первичная фотохимическая реакция фотосинтеза состоит в диссоциации воды, а не в разложении С02. Способные к фотосинтетической ассимиляции С02 бактерии (за исключением цианобактерий) нуждаются в восстановителях типа H2S, Н2, СН3 и других и не выделяют в процессе фотосинтеза кислород. Такой тип фотосинтеза был назван фоторедукцией. Ван Ниль пришел к выводу, что для пурпурных или зеленых серобактерий общее уравнение фотосинтеза может быть представлено следующим образом: С02 + H2A (СН20) + H2O + 2A
Голландский микробиолог К.Б. ван Ниль, изучая особенности бактериального фотосинтеза и сравнивая его с фотосинтезом у растений, в 1937—1941 гг. также пришел к заключению, что первичная фотохимическая реакция фотосинтеза состоит в диссоциации воды, а не в разложении С02. Способные к фотосинтетической ассимиляции С02 бактерии (за исключением цианобактерий) нуждаются в восстановителях типа H2S, Н2, СН3 и других и не выделяют в процессе фотосинтеза кислород. Такой тип фотосинтеза был назван фоторедукцией. Ван Ниль пришел к выводу, что для пурпурных или зеленых серобактерий общее уравнение фотосинтеза может быть представлено следующим образом: С02 + H2A (СН20) + H2O + 2A
![]() где Н2А — окисляемый субстрат (донор водорода). Он предположил, что для высших растений и водорослей Н2А — это Н20, а 2А — это 02. Тогда первичным фотохимическим актом в фотосинтезе растений должно быть разложение воды на окислитель (ОН) и восстановитель (Н). Затем первичный восстановитель (Н) восстанавливает С02, а первичный окислитель (ОН) участвует в реакции, в которой высвобождается 02 и снова образуется H20. B соответствии с этим предположением полное уравнение фотосинтеза, по ван Нилю, можно записать так: CO2 + 4H2O хлорофилл (CH2O) + 3H2O + O2 , причем эта суммарная реакцияtслагается из трех этапов:
где Н2А — окисляемый субстрат (донор водорода). Он предположил, что для высших растений и водорослей Н2А — это Н20, а 2А — это 02. Тогда первичным фотохимическим актом в фотосинтезе растений должно быть разложение воды на окислитель (ОН) и восстановитель (Н). Затем первичный восстановитель (Н) восстанавливает С02, а первичный окислитель (ОН) участвует в реакции, в которой высвобождается 02 и снова образуется H20. B соответствии с этим предположением полное уравнение фотосинтеза, по ван Нилю, можно записать так: CO2 + 4H2O хлорофилл (CH2O) + 3H2O + O2 , причем эта суммарная реакцияtслагается из трех этапов:
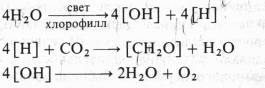
Идеи ван Ниля были поддержаны результатами опытов английского физиолога растений Р. Хилла, который в 1937 г. показал, что изолированные хлоропласты под действием света способны разлагать воду и выделять кислород в присутствии акцепторов электронов (феррицианида, бензохинона и др.). Это явление получило название реакции Хилла. Согласно Хиллу, процесс разложения воды осуществляется в три этапа:
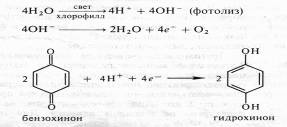
Прямые экспериментальные доказательства того, что кислород при фотосинтезе освобождается именно из воды, были получены в 1941 г. независимо в СССР и в США. А.П.Виноградов и Р.В.Тейс с помощью масс-спектрометра показали, что отношение 160:180 в кислороде, выделяющемся при фотосинтезе, соответствует соотношению этих изотопов в воде, а не в диоксиде углерода. Группа американских ученых, возглавляемая С.Рубеном и М.Каменом, одновременно проводила опыты с водорослями. В одной камере, куда помещались водоросли, вода содержала тяжелый изотоп кислорода (Н2180), а кислород С02 не метился. Во второй камере, наоборот, использовался меченый С02 (С1802), а в состав воды входил обычный кислород. При освещении водорослей в этих камерах 1802 выделялся в значительно большем количестве, если метка была у кислорода воды.
2 Фотосинтез как основа энергетики биосферы. Космическая роль фотосинтеза. Роль фотосинтеза в процессах энергетического и пластического обмена растительного организма
К.А.Тимирязев, который первым начал изучать космическую роль зеленых растений, в публичной лекции, прочитанной в 1875 г., следующим образом представил эту проблему слушателям: «...луч солнца... упал на зеленую былинку пшеничного ростка ... Он ... затратился на внутреннюю работу ... превратясь в растворимый сахар ... отложился, наконец, в зерне в виде крахмала или в виде клейковины. В той или другой форме он вошел в состав хлеба, который послужил нам пищей. Он преобразился в наши мускулы, в наши нервы. Этот луч солнца согревает нас. Он приводит нас в движение. Быть может, в эту минуту он играет в нашем мозгу».
Действительно, фотосинтез — единственный процесс на 3емле, идущий в грандиозных масштабах и связанный с превращением энергии солнечного света в энергию химических связей. Эта космическая энергия, запасенная зелеными растениями, составляет основу для жизнедеятельности всех других гетеротрофных организмов на Земле от бактерий до человека. Выделяют пять аспектов космической и планетарной роли растений, которые рассмотрены ниже:
1. Накопление органической массы. В процессе фотосинтеза наземные растения образуют 100—172 млрд. т, а растения морей и океанов — 60 — 70 млрд. т биомассы в год (в пересчете на сухое вещество). Общая масса растений на Земле в настоящее время составляет 2402,7 млрд. т, причем 90% этой сухой массы приходится на целлюлозу. На долю наземных растений приходится 2402,5 млрд. т, а на растения гидросферы — всего 0,2 млрд. т (из-за недостатка света). Общая масса, животных и микроорганизмов на Земле — 23 млрд. т, что составляет около 1 % от растительной биомассы. Из этого количества 20 млрд. т приходится на обитателей суши, а 3 млрд. т — на животных и микроорганизмы гидросферы.
За время существования жизни на Земле органические остатки растений и животных накапливались и модифицировались. На суше эти органические вещества представлены в виде подстилки, гумуса и торфа, из которых, при определенных условиях, в толще литосферы формировался уголь. В морях и океанах органические остатки (главным образом животного происхождения) оседали на дно и входили в состав осадочных пород. При опускании в более глубокие области литосферы из этих остатков под действием микроорганизмов, повышенных температур и давления образовывались газ и нефть. Масса органических веществ подстилки, торфа и гумуса оценивается в 194, 220 и 2500 млрд. т соответственно. Нефть и газ составляют 10000-12000 млрд. т. Содержание органических веществ в осадочных породах достигает 20 000 000 млрд. т (по углероду).
Особенно интенсивное накопление мертвых органических остатков происходило 300 млн. лет назад в палеозойскую эру. Запасы древесины, а в последние 200 лет угля, нефти и газа используются человеком для получения энергии, необходимой в быту, промышленности и сельском хозяйстве.
2. Обеспечение постоянства содержания CO2 в атмосфере. Образование органических веществ гумуса, осадочных пород и горючих ископаемых выводило значительные количества С02 из круговорота углерода. В атмосфере Земли С02 становилось все меньше и в настоящее время он составляет только 0,03% (по объему), или (в абсолютных значениях) 711 млрд. т в пересчете на углерод.
В кайнозойской эре содержание диоксида углерода в атмосфере стабилизировалось и испытывало лишь суточные, сезонные и более длительные геохимические колебания. Эта стабилизация достигается сбалансированным связыванием и освобождением С02, осуществляемых в глобальном масштабе. Связывание С02 в ходе фотосинтеза и образование карбонатов компенсируется выделением С02 за счет других процессов. Ежегодное поступление С02 в атмосферу в пересчете на углерод (в млрд. т) обусловлено: дыханием растений — 10, дыханием и брожением микроорганизмов — 25, дыханием животных и человека — 1,6, производственной деятельностью людей — 5. геохимическими процессами — 0.05. При отсутствии этого поступления весь С02 атмосферы был бы связан в ходе фотосинтеза за 6 — 7 лет. Мощным резервом диоксида углерода является Мировой океан, в водах которого растворено в 60 раз больше С02, чем находится в атмосфере. Фотосинтез, с одной стороны, дыхание организмов и карбонатная система океана, с другой, поддерживают относительно постоянный уровень С02 в атмосфере.
Однако за последние десятилетия из-за все более возрастающего сжигания человеком горючих ископаемых, а также из-за вырубки лесов и разложения гумуса содержание С02 в атмосфере начало увеличиваться примерно на 0,23% в год. Это обстоятельство может иметь далеко идущие последствия в связи с тем, что концентрация С02 оказывает влияние на тепловой режим Земли.
3. Парниковый эффект. Поверхность Земли получает теплоту главным образом от Солнца. Часть этой теплоты поступает обратно в космос в виде инфракрасных лучей. Диоксид углерода в атмосфере, а также вода поглощают инфракрасное излучение и таким образом сохраняют значительное количество теплоты на Земле (парниковый эффект). Микроорганизмы и растения в процессе дыхания или брожения поставляют 85 % общего количества С02, поступающего ежегодно в атмосферу, и вследствие этого оказывают влияние на тепловой режим нашей планеты.
Тенденция к повышению содержания С02 в атмосфере из-за сжигания огромных количеств нефти, газа и из-за других причин, указанных выше, может способствовать увеличению средней температуры на поверхности Земли, что приведет к ускорению таяния ледников в горах и на полюсах, затоплению прибрежных зон. Возможно, однако, что повышение концентрации С02 будет способствовать усилению фотосинтеза растений, что устранит избыточное накопление диоксида углерода. Известно, что изменение концентрации С02 в биосфере выступает как элемент обратной связи.
4. Накопление кислорода в атмосфере.
Первоначально в атмосфере Земли 02 присутствовал в следовых количествах. В настоящее время он составляет 21 % по объему воздуха. Появление и накопление 02 в атмосфере связано с жизнедеятельностью зеленых растений. Ежегодно в ходе фотосинтеза кислород поступает в атмосферу в количестве 70— 120 млрд. т. Этот кислород необходим для дыхания всех гетеротрофов — бактерий, грибов, животных и человека, а также зеленых растений в ночное время. Особое значение в поддержании высокой концентрации 02 в атмосфере имеют леса. Подсчитано, что 1 га леса весной и летом за час выделяет 02 в количестве, достаточном для дыхания 200 человек.
5.Озоновый экран. Еще одно важнейшее следствие выделения растениями кислорода — образование озонового экрана в верхних слоях атмосферы на высоте около 25 км. Озон (03) образуется в результате фотодиссоциации молекул 02 под действием солнечной радиации. Озон задерживает большую часть ультрафиолетовых лучей (240 — 290 нм), губительно действующих на все живое. Возможность частичного разрушения озонового экрана из-за загрязнения атмосферы промышленными и другими отходами — серьезная проблема охраны биосферы.
3 Структурная организация фотосинтетического аппарата
3.1 Лист как орган фотосинтеза
Основные функции листа зеленого растения — фотосинтез (воздушное питание), транспирация (регулируемое испарение воды) и синтез ряда органических веществ, в том числе некоторых фитогормонов (ауксина, гиббереллинов, абсцизовой кислоты).
Лист, как правило, имеет плоскую форму и дорсовентральное строение. Пластинчатая форма листа обеспечивает наибольшую поверхность на единицу объема ткани, что создает наилучшие условия для воздушного питания.
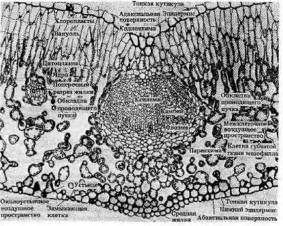
Поперечный разрез листовой пластинки и средней жилки листа Бирючины – типичного двудольного растения
Важнейшая ткань листа — мезофилл, где осуществляется фотосинтез. Покрывающий лист эпидермис, клетки которого, за исключением замыкающих клеток устьиц, не содержат хлоропластов, защищает ткани листа, регулирует газообмен и транспирацию. Система разветвленных проводящих пучков необходима для снабжения тканей листа водой, минеральными и некоторыми органическими веществами и для оттока ассимилятов в другие части растения. Мезофилл обычно дифференцирован на две ткани — палисадную (столбчатую), расположенную под верхним эпидермисом, и губчатую, находящуюся в нижней стороне листа. В палисадном мезофилле клетки вытянуты перпендикулярно поверхности листа и расположены в один или несколько слоев. Клетки губчатого мезофилла связаны друг с другом более рыхло из-за больших межклетников. У большинства растений устьица находятся нанижней стороне листа, обширные межклетники губчатой паренхимы обеспечивают газообмен. Благодаря развитой системе межклетников мезофилл обладает громадной поверхностью, во много раз превышающей наружную поверхность листа.
Палисадная ткань, обращенная к свету, содержит большую часть всех хлоропластов листа и выполняет основную работу по ассимиляции С02. Число слоев клеток мезофилла в листе в большой степени зависит от интенсивности освещения. Листья, выросшие на ярком свету, имеют как правило, более развитый столбчатый мезофилл. У растений засушливых ареалов — ксерофитов — палисадная паренхима часто расположена по обеим сторонам листа, а губчатая сильно редуцирована или совсем отсутствует.
3.2Состав, строение, функции, возникновение и развитие хлоропластов
Хлоропласты имеют зеленый цвет, обусловленный присутствием основного пигмента — хлорофилла. Хлоропласты содержат также вспомогательные пигменты — каротиноиды (оранжевого цвета). По форме хлоропласты — это овальные линзовидные тельца размером (5—10) * (2—4) мкм. В одной клетке листа может находиться 15—20 и более хлоропластов, а у некоторых водорослей — лишь 1-2 гигантских хлоропласта (хроматофора) различной формы.
Хлоропласты ограничены двумя мембранами — наружной и внутренней. Наружная мембрана отграничивает жидкую внутреннюю гомогенную среду хлоропласта — строму (матрикс). В строме содержатся белки, липиды, ДНК (кольцевая молекула), РНК, рибосомы и запасные вещества (липиды, крахмальные и белковые зерна) а также ферменты, участвующие в фиксации углекислого газа.
Внутренняя мембрана хлоропласта образует впячивания внутрь стромы — тилакоиды, или ламеллы, которые имеют форму уплощенных мешочков (цистерн). Несколько таких тилакоидов, лежащих друг над другом, образуют грану, и в этом случае они называются тилакоидами граны. Именно в мембранах тилакоидов локализованы светочувствительные пигменты, а также переносчики электронов и протонов, которые участвуют в поглощении и преобразовании энергии света.
Хлоропласты в клетке осуществляют процесс фотосинтеза.
Лейкопласты — мелкие бесцветные пластиды различной формы. Они бывают шаровидными, эллипсоидными, гантелевидными, чашевидными и т. д. По сравнению с хлоропластами у них слабо развита внутренняя мембранная система. Лейкопласты в основном встречаются в клетках органов, скрытых от солнечного света (корней, корневищ, клубней, семян). Они осуществляют вторичный синтез и накопление запасных питательных веществ — крахмала, реже жиров и белков.
Хромопласты отличаются от других пластид своеобразной формой (дисковидной, зубчатой, серповидной, треугольной, ромбической и др.) и окраской (оранжевые, желтые, красные). Хромопласты лишены хлорофилла и поэтому не способны к фотосинтезу. Внутренняя мембранная структура их слабо выражена.
Хромопласты присутствуют в клетках лепестков многих растений (лютиков, калужниц, нарциссов, одуванчиков и др.), зрелых плодов (томаты, рябина, ландыш, шиповник) и корнеплодов (морковь, свекла), а также листьев в осеннюю пору. Яркий цвет этих органов обусловлен различными пигментами, относящимися к группе каргиноидов, которые сосредоточены в хромопластах.
4 Пигменты хлоропластов
Пигменты — важнейший компонент аппарата фотосинтеза. Изучение растительных пигментов резко ускорилось благодаря работам русского физиолога растений М. С. Цвета. Пытаясь найти способ разделения пигментов на индивидуальные вещества, Цвет в 1901 — 1903 гг. открыл принципиально новый метод, который он назвал адсорбционной хроматографией. Через колонку с сорбентом пропускается растворитель с растворенными веществами. Так как вещества различаются по степени адсорбции, они перемещаются по колонке с разной скоростью. В результате происходит разделение веществ. Этот прием широко используется в современной биохимии, химии и в некоторых отраслях промышленности. С помощью хроматографического метода Цвет обнаружил два хлорофилла — а и и разделил желтые пигменты листа на три фракции.
Пигменты пластид относятся к трем классам веществ: хлорофиллам, фикобилинам и каротиноидам.
4.1 Хлорофиллы: состав, структура, биосинтез, оптические и химические свойства, значение
Впервые хлорофилл в кристаллическом виде был описан русским физиологом и ботаником И. П. Бородиным в 1883 г. В дальнейшем оказалось, что это не сам хлорофилл, а несколько видоизмененная его форма — этилхлорофиллид.
Польские биохимики М. Ненцкий и Л. Мархлевский (1897) обнаружили, что основу молекулы хлорофилла, как и гема гемоглобина, составляет порфириновое кольцо. Таким образом было показано принципиальное структурное сходство этих пигментов у растений и животных.
Немецкий химик Р. Вильштеттер в 1906—1914 гг. установил элементарный состав хлорофилла а — C55H72O5N4Mg и хлорофилла b — C55H70O6N4Mg, а немецкий биохимик Г. Фишер в 1930—1940 гг. полностью расшифровал структурную формулу хлорофилла. В 1960 г. химики-органики Р.Б. Вудворд (США) и М. Штрель (ФРГ) осуществили искусственный синтез хлорофилла. Хлорофилл — сложный эфир дикарбоновой кислоты хлорофиллина, у которой одна карбоксильная группа этерифицирована остатком метилового спирта, а другая остатком одноатомного непредельного спирта фитола. Структурная формула хлорофилла а представлена на рисунке:
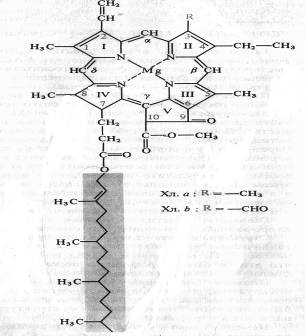
Четыре пиррольных кольца (I — IV) соединены между собой метановыми мостиками, образуя порфириновое ядро. Наружные атомы углерода в тетрапиррольном цикле обозначены цифрами 1 — 10. Атомы азота пиррольных колец четырьмя координационными связями взаимодействуют с атомом магния. В структуре порфиринового ядра есть также циклопентановое кольцо (V), образованное остатком кетопропионовой кислоты и содержащее химически активную карбонильную группу у С9 и метилированную карбоксильную группу у С10. Структура, состоящая из тетрапиррольного и циклопентанового колец, получила название форбина. Боковая цепь IV пиррольного кольца включает в себя пропионовую кислоту, связанную сложноэфирной связью с полиизопреновым непредельным спиртом фитолом (С20Н39ОН). У 1, 3, 5 и 8-го углеродов пиррольных колец имеются метальные группы, у 2-го — винильная, у 4-го — этильная группа. Порфириновое кольцо представляет собой систему из девяти пар конъюгированных (сопряженных) чередующихся двойных и одинарных связей с 18 делокализованными p-электронами. Хлорофилл b отличается от хлорофилла а тем, что у 3-го углерода вместо метильной находится формильная (—СНО) группа. Структура хлорофилла, лишенная фитола, называется хлорофиллидом. При замещении атома магния протонами в молекуле хлорофиллов образуются соответствующие феофетины.
У высших растений и водорослей обнаружены хлорофиллы а, b, с, d. Все фотосинтезирующие растения, включая все группы водорослей, а также цианобактерии, содержат хлорофиллы группы а. Хлорофилл b представлен у высших растений, у зеленых водорослей и эвгленовых. У бурых и диатомовых водорослей вместо хлорофилла b присутствует хлорофилл c, а у многих красных водорослей — хлорофилл d. Вфотосинтезирующих бактериях, осуществляющих фоторедукцию, найдены различные бактериохлорофиллы.
Биосинтез хлорофиллов. Первый этап биосинтеза хлорофиллов у растений — образование 5-аминолевулиновой кислоты (А Л К) из С5-дикарбоновых кислот. Показано, что глутаминовая кислота через 2-гидроксиглутаровую превращается в 4,5-диоксовалериановую, которая затем аминируется за счет аланина или других аминокислот (рис. 5.2). Реакция переаминирования катализируется АЛК-трансаминазой с участием пиридоксальфосфата в качестве кофермента. Для синтеза АЛК может использоваться и оскетоглутаровая (2-оксоглутаровая) кислота. Циклизация двух молекул АЛК приводит к образованию пиррольного соединения — порфобилиногена. Из четырех пиррольных колец формируется уропорфириноген, который превращается в протопорфирин IX. Дальнейший путь превращений протопорфирина может быть различным. С участием железа формируется гем, входящий в состав цитохромов, каталазы, пероксидазы и гемоглобина. Если молекулу протопорфирина включается магний, затем карбоксильная группа у С10 этерифицируется метильной группой 6-аденозил-b-метионина и замыкается циклопентановое кольцо (К), то образуется протохлорофиллид. Под действием света в течение нескольких секунд протохлорофиллид превращается в хлорофиллид а в результате гидрирования двойной связи у С7 — С8 в IV пиррольном ядре. У низших растений и некоторых голосеменных (у хвойных) хлорофиллид может образовываться в темноте. Хлорофиллид обладает теми же спектральными свойствами, что и хлорофилл. Последний этап в формировании молекулы хлорофилла а — этерификация фитолом, который, как и все полиизопреновые соединения, синтезируется из ацетил-KоА через мевалоновую кислоту. По данным А. А. Шлыка (1965), хлорофилл b может образовываться из вновь синтезированных молекул хлорофилла а. Все описанные процессы, начиная с образования АЛК, осуществляются в хлоропластах. Первые признаки зеленения, выращенных в темноте этиолированных покрытосеменных растений, наблюдаются через 2 — 4 ч после начала освещения.
Физико-химические свойства хлорофиллов.
В твердом виде хлорофилл а представляет собой аморфное вещество сине-черного цвета. Температура плавления хлорофилла а 117 — 120 °С. Хлорофиллы хорошо растворимы в этиловом эфире, бензоле, хлороформе, ацетоне, этиловом спирте, плохо растворимы в петролейном эфире и нерастворимы в воде. Раствор хлорофилла а в этиловом эфире имеет сине-зеленый цвет, хлорофилла b — желто-зеленый. Резко выраженные максимумы поглощения хлорофиллов лежат в красной и синей частях спектра.
В этиловом эфире максимумы поглощения хлорофиллов группы а в красной части спектра — в пределах 660 — 663 нм, в синей — 428 — 430 нм, хлорофилла b — соответственно в пределах 642 — 644 и 452 — 455 нм. Хлорофиллы очень слабопоглощают оранжевый и желтый свет и совсем не поглощают зеленые и инфракрасные лучи. Поглощение в сине-фиолетовой части спектра обусловлено системой конъюгированных одинарных и двойных связей порфиринового кольца молекулы хлорофилла. Интенсивная полоса поглощения в красной области связана с гидрированием двойной связи у С7 — С8 в IV пиррольном ядре (при переходе от протохлорофиллида к хлорофиллиду) и присутствием магния в порфириновом кольце. Эти же условия способствуют снижению поглощения в желтой и зеленой частях спектра. Замещение магния протонами при обработке хлорофилла кислотой приводит к образованиюфеофетина, имеющего буро-зеленый цвет и ослабленный красный максимум поглощения. Удаление остатков фитола и метилового спирта путем щелочного гидролиза мало сказывается на спектре поглощения хлорофилла. У хлорофиллина те же спектральные свойства, что и у хлорофилла: он зеленый, имеет красный и синий максимумы поглощения.
На положение максимумов спектра поглощения оказывают влияние природа растворителя и взаимодействие молекул хлорофилла друг с другом, а также с другими пигментами, липидами и белками. У агрегированных молекул хлорофилла (например, в твердых пленках и у хлорофилла, находящегося в хлоропластах) красный максимум поглощения сдвинут в более длинноволновую область (до 680 нм).
Растворы хлорофиллов в полярных растворителях обладают яркой флуоресценцией (люминесценцией). В этиловом эфире у хлорофилла а наблюдается рубиново-красная флуоресценция с максимумом 668 нм, у хлорофилла b — 648 нм т. е. максимумы флуоресценции в соответствии с правилом Стокса несколько сдвинуты в более длинноволновую часть спектра по отношению к максимумам поглощения. Агрегированный хлорофилл и хлорофилл в нативном состоянии (в живом листе) флуоресцируют слабо. Растворы хлорофиллов способны также к фосфоресценции (т. е. длительному послесвечению), максимум которого лежит в инфракрасной области.
Механизмы флуоресценции и фосфоресценции, в том числе молекул хлорофилла, достаточно хорошо изучены. Наиболее устойчивы те состояния атомов и молекул, в которых валентные электроны занимают самые низкие энергетические уровни и распределены по ним согласно принципу Паули (не более двух электронов с антипараллельными спинами на каждой орбитали). Такое состояние молекулы называют основным синглетным (S0) энергетическим состоянием. Суммарный спин (вектор магнитных моментов) всех электронов молекулы в этом случае равен нулю. Когда молекула поглощает квант света, то за счет этой энергии электрон переходит на более высокие вакантные орбитали. Если у возбужденного электрона сохраняется то же направление спина, то молекула находится в синглетном возбужденном состоянии (S*). Если при переходе на более высокую орбиталь спин электрона обращается, то такое возбуждение называется триплетным (T*). Прямой переход из основного состояния в триплетное — маловероятное событие, так как для этого необходимо обращение ориентации спина. Переходу молекулы из основного состояния в возбужденное, происходящему при поглощении кванта света, соответствует полоса в спектре поглощения.
Поглощение молекулой хлорофилла кванта красного света приводит к синглетному электронвозбужденному состоянию — S*1.
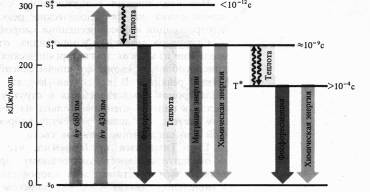
При поглощении кванта синего света с более высоким уровнем энергии электрон переходит на более высокую орбиту (S*2). Возбужденная молекула хлорофилл