


Психология и физиология сна
Качество нашей жизни в значительной степени определяется состоянием умственного и физического здоровья. Общество осознало, насколько важны для сохранения здоровья правильное питание и физическая активность. Значение же снаво многом остается недооцененным. Это обусловлено тем, что мы анализируем свои ощущения и происходящие с нами события во время бодрствования и рассматриваем сон, как пассивный период отдыха для души и тела.
Однако, необходимо задуматься:
· Природа настолько наивна, что сохранила сон в процессе эволюции и отобрала у людей и животных треть их жизни?
· Почему стоит не выспаться одну ночь — и уже болит голова, плохое настроение, снижена работоспособность?
Современная наука убедительно показала, что сон— это активный, сложный и многофункциональный процесс. Около 25% периода сна мозг настолько же активен, как при бодрствовании. Он проявляет высокую электрическую активность и ему требуется интенсивный приток крови. Сон важен для восстановления энергетических затрат мозга и тела, для нормального функционирования памяти. Во время сна вырабатываются определенные биологически активные вещества, как, например, гормон роста. В период сна усиливается деление клеток и происходит множество других процессов. Если в эксперименте животное лишить сна, то оно погибает.
Изучение сна привело к выявлению большого количества нарушений сна и выделению нового раздела медицины — медицины сна или, как ее еще называют, сомнологии — науки о сне. Диагностика и лечение различных расстройств сна требует не только специальных врачебных знаний, но и определенного, отвечающего современным требованиям, медицинского оборудования.
Я считаю тему о природе и тайнах сна очень интересной и познавательной и решила узнать о сомнологии как можно больше. Итак, тема моего реферата «СОМНОЛОГИЯ».
Сон родился в тепле
Считается, что сон появился вместе с терморегуляцией. Большая часть млекопитающих приспособилась к жизни путем организации терморегуляции (некоторые исследователи связывают это с великим оледенением).

В отличие от холоднокровных животных, у которых сна нет, а есть лишь бодрствование и покой, во время которого мозг отключается, мозг теплокровных активен и в состоянии покоя. В результате эволюции появилось особое состояние — сон; не покой и не бодрствование — во сне мозг работает. Иначе, чем в состоянии бодрствования, но почти так же активно.
Объяснить природу сна пытались с самых давних времен. Это еще до нашей эры пытались сделать такие мощные умы, как Эмпедокл, Гиппократ, Аристотель. Эмпедокл полагал, что сон возникает из-за отделения огня от остальных первоэлементов — воздуха, воды и земли, а попросту из-за уменьшения тепла в крови. «Отец медицины» Гиппократ изъяснялся проще, но, по сути, похоже: сон приходит из-за оттока крови и тепла во внутренние органы. Аристотель называл тепло «духами», которые появляются в результате поедания пищи и вызывают сонливость. В XVI веке Парацельс вполне приземленно объяснял причину сна дневным утомлением и призывал ложиться с закатом и вставать с рассветом. Было еще немало объяснений все с теми же мистическими терминами вроде «нервных духов» или «жизненных эфиров». Самым симпатичным мне показалось высказанное уже в XIX веке философское определение профессора физиологии и хирургии Филиппа Франца фон Вальтера: «Сон — это капитуляция нашей эгоистичной сущности перед коллективизмом естественного духа, слияние индивидуальной человеческой души с универсальным духом природы».
В пору расцвета естественных наук появились более рациональные объяснения загадки сна — изменения в нервных клетках, обеднение мозга кислородом и прочее. Однако проверить все эти гипотезы не было никакой возможности. Да и как проверять, если у объекта изучения не спросишь, что с ним происходит. Он может уже после сна лишь попытаться описать его качество — плохо или хорошо спал, просыпался ли, снилось ли что, а если снилось, то, что именно. В результате можно было лишь дать некие внешние характеристики вроде закрытых глаз, замедления дыхания, пульса, возможно, некоторого снижения температуры. Но эти же характеристики были вполне уместны при описании просто отдыхающего с закрытыми глазами человека.
Каждую ночь мы спим, и, как ни приятно нам это занятие, мы не понимаем, что мы делаем. Назначение сна все еще остается загадкой.
Сон – периодически наступающее физиологическое состояние у человека и у животных. Он характеризуется почти полным отсутствием реакций на внешние раздражения и уменьшением активности ряда физиологических процессов. Различают нормальный (физиологический) сон и несколько видов патологического сна (наркотический, летаргический и другие).
Метод изучения сна - полисомнография
Сон можно определить как «особое генетически детерминированное состояние организма человека (и теплокровных животных, т.е. млекопитающих и птиц), характеризующееся закономерной последовательной сменой определенных полиграфических картин в виде циклов, фаз и стадий» (В.М. Ковальзон).
Это сложное определение сна свидетельствует, во-первых, что сон - это неотъемлемая часть бытия человека, т.к. необходимость спать генетически предопределена и, во-вторых, что состояние сна может быть определено только с помощью специальных объективных методов. Для объективного изучения сна и его нарушений используется полисомнография (ПСГ). Это основной метод исследования сна человека.
Полисомнография - метод регистрации показателей жизнедеятельности во время сна. Слово происходит от слов poly- множественный, somnos - сон, grapho - пишу. Обычно ПСГ производится во время ночного сна.
Полисомнография - метод, включающий параллельную регистрацию электроэнцефалограммы (ЭЭГ) (как правило, многоканальной), электроокулограммы (ЭОГ), электромиограммы (ЭМГ), электрокардиограммы (ЭКГ), артериального давления (АД), двигательной активности (общей и в конечностях), дыхательных движений грудной клетки и брюшной стенки, оро-назального тока воздуха, уровня сатурации кислорода в крови, выраженности храпа, температуры тела, видеомониторирование и т.д.
Целью проведения полисомнографического исследования является объективизация деятельности организма человека во время сна. Для этого регистрируется ряд показателей, наиболее важными из которых являются следующие:
Электроэнцефалограмма (ЭЭГ) - запись электрической активности мозга
Электроокулограмма (ЭОГ) - запись движений глаз
Электромиограмма (ЭМГ) - запись напряжения мышц (чаще подбородочных).
Эти три показателя (ЭЭГ, ЭОГ и ЭМГ) являются базовыми для идентификации стадий и фаз сна. На основании анализа этих показателей строится гипнограмма, отражающая динамику стадий и фаз сна в течение периода регистрации.
Совокупность этих трех показателей позволяет в любой момент времени определить, в какой стадии сна находится человек. По результатам записи сна строится временной график сна - гипнограмма, а затем вычисляются различные характеристики сна (время засыпания, продолжительность стадий сна и т.д.) и сравниваются с нормативными. ПСГ позволяет ответить на два основных вопроса:
1. Соответствуют ли показатели сна пациента нормативным? (при этом производится визуальный анализ гипнограммы, а количественные данные сравниваются с возрастной нормой).
2. Какие феномены встречаются во время сна и могут быть причиной его нарушения? (например: апноэ, мышечные подергивания, эпилептические разряды). Для того, чтобы получить данные не только о структуре сна, но и о вредных феноменах, связанных со сном, при ПСГ регистрируется еще несколько показателей:
Поток выдыхаемого воздуха изо рта и ноздрей
Дыхательные движения груди и живота
Шум дыхания (храп)
Уровень насыщения крови кислородом
Положение тела в постели
Число сердечных сокращений
Полисомнографическое исследование проводится в Центре сна, в специально оборудованной одноместной палате. Вечером пациент приходит в Центр, заполняет специальную анкету и готовится ко сну. Затем на него накладывают (наклеивают) датчики, он ложится в кровать и спит. В течение всей ночи показатели его сна записываются на компьютер и видеомагнитофон, а сотрудник Центра наблюдает за его состоянием. Утром с пациента снимают датчики и сообщают, когда приходить за результатами.
Есть и другие варианты полисомнографии.
Для диагностики храпа и апноэ во сне могут использоваться упрощенные варианты полисомнографии - ночное и дневное кардиореспираторное мониторирование. При этом накладываются только датчики для оценки дыхания во сне. Время сна определяется по данным видеомониторирования. В дневное время при трудностях засыпания пациент получает короткодействующий снотворный препарат, не изменяющий картину дыхания во сне.
Для количественной оценки дневной сонливости используется множественный тест латенции сна (МТЛС). В течение дневного времени 5 раз пациента укладывают спать с наложенными как при ПСГ датчиками. Затем усредняют время засыпания и сравнивают с нормативным. Этот тест используется для диагностики нарколепсии.
Сон человека представляет собой континуум (последовательность) функциональных состояний мозга - 1, 2, 3 и 4 стадии фазы медленного сна (ФМС) и фазу быстрого сна (ФБС).
Стадии ФМС и ФБС составляют один цикл сна, и таких циклов у здорового человека бывает от 4 до 6 за ночь; эти циклы не одинаковы: в первых двух максимально представлена ФМС, а в утренних - ФБС.
Основная функция ФМС - восстановительная (накопление энергетических, в первую очередь, фосфатергических связей, синтез пептидов и нуклеиновых кислот, в этой фазе наблюдаются пики секреции соматотропного гормона, пролактина, мелатонина, а ФБС - переработка информации и построение программы поведения.
При физиологическом старении организма структура сна претерпевает следующие изменения: снижается общая длительность сна, увеличивается длительность поверхностных стадий, увеличивается время засыпания и время бодрствования внутри сна, возрастает двигательная активность во сне, возникает фрагментация сна, отмечается большая сохранность ФБС по отношению к ФМС, выявляется полифазность сна (наличие дневного и ночного сна) и дневные микрозасыпания. Жалобы на плохой ночной сон у людей старше 60 лет встречается в 3-4 раза чаще, чем в среднем возрасте.
Один из главных вопросов, волновавших физиологов еще со времен И.П.Павлова - это существование в мозге «центра сна». Прямое изучение нейронов, вовлеченных в регуляцию сна-бодрствования, показало, что нормальное функционирование таламо-кортикальной системы мозга, обеспечивающее весь спектр сознательной деятельности человека в бодрствовании, возможно только при наличии мощных воздействий со стороны структур, называемых активирующими. Благодаря этим воздействиям мембрана значительного большинства корковых нейронов в бодрствовании деполяризована и только в таком состоянии деполяризации эти нейроны способны обрабатывать и отвечать на сигналы, приходящие к ним от других нервных клеток. Таких систем активации мозга (их можно условно назвать «центрами бодрствования»), вероятно, пять или шесть, и они локализуются на всех церебральных уровнях: в ретикулярной формации ствола, в области синего пятна и дорсальных ядер шва, в заднем гипоталамусе и базальных ядрах переднего мозга. У человека нарушение деятельности любой из этих систем не может быть скомпенсировано за счет других, несовместимо с сознанием и приводит к коме.
Казалось логичным предположить, что если в мозге есть «центры бодрствования», то должны быть и «центры сна». Однако детальное изучение нейронов показало, что в систему поддержания бодрствования встроены механизмы положительной обратной связи, в виде особых нейронов, функцией которых является торможение активирующих нейронов, и которые сами тормозятся этими нейронами. Эти нейроны разбросаны по разным отделам мозга, хотя и отмечено их скопление в ретикулярной части черного вещества; общим для них является выделение одного и того же химического посредника - гамма-аминомасляной кислоты, главного тормозного вещества мозга. Как только активирующие нейроны ослабляют свою активность, включаются тормозные нейроны и ослабляют ее еще более. Процесс развивается по нисходящей в течение некоторого времени, пока не срабатывает некий «триггер» и вся система перебрасывается в другое состояние - бодрствования или парадоксального сна. Отражением этого процесса является смена картин в электрической активности головного мозга по ходу 90-минутного цикла сна человека.
Еще одна эволюционно древняя тормозная система головного мозга использует в качестве медиатора аденозин. Становится очевидной роль этой системы в формировании гиперсомнии (при черепно-мозговой травме, менингите, африканской "сонной болезни" и других заболеваниях).
В экспериментах на лабораторных животных показано, что, по мере углубления сна доминируют все более мощные тормозные постсинаптические потенциалы, перемежающиеся периодами активации - по типу «пачка-пауза». В этих условиях ухудшается возможность переработки информации в мозге. Разряды нейронных активирующих систем прогрессивно урежаются. Таким образом, в медленном сне происходит восстановление мозгового гомеостаза и другие восстановительные процессы, например синтез фосфатергических соединений («накопители энергии»), гормона роста (соматотропного гормона), белков и нуклеиновых кислот. С этой точки зрения бодрствование и медленный сон - как бы «две стороны одной медали». Отсутствие единого «центра медленного сна» (с учетом его значимости) делает систему его организации более надежной, не зависящей полностью от деятельности одного центра в случае каких-либо нарушений его функционирования. Вместе с тем длительное тотальное подавление медленного сна невозможно, так как он должен периодически сменять бодрствование и в условиях искусственного подавления сна мозг идет на различные уловки, лишь бы сохранить представленность медленного сна. Важно также и то, что в условиях медленного сна переработка информации мозгом не прекращается, а изменяется: от обработки экстероцептивных (внешних) мозг переходит к обработке интероцептивных (внутренних) импульсов. Таким образом, функция медленного сна, включает не только восстановительные процессы, но и оптимизацию управления внутренними органами.
В отличие от медленно сна быстрый (парадоксальный) сон запускается из определенного центра, расположенного в задней части мозга, в области варолиева моста и продолговатого мозга. Медиаторами этих клеток служат ацетилхолин, глутаминовая и аспарагиновая кислоты. Во время быстрого сна клетки мозга активны, однако информация от органов чувств (афферентная) к ним не поступает, и нисходящая (эфферентная) на мышечную систему не подается. В этом и заключается парадоксальный характер этого состояния. При этом интенсивно перерабатывается та информация, которая была получена в предшествующем бодрствовании и хранится в памяти; кроме того, в быстром сне происходит формирование будущей программы поведения. Неадекватные включения «центра парадоксального сна» действительно имеют место при некоторых, довольно редких видах генетически обусловленной патологии (нарколепсия и др.). В отличие от бодрствования в быстром сне функционируют лишь активирующие системы, локализованные в ретикулярной формации ствола и использующие в качестве передатчиков ацетилхолин, глутаминовую и аспарагиновую кислоты. Все же остальные активирующие системы выключаются, и их нейроны неактивны весь период парадоксального сна. Это молчание значительного количества активирующих систем мозга и является тем фундаментальным фактом, который определяет различие между бодрствованием и парадоксальным сном на физиологическом уровне.
Традиционно обсуждаемые нейрохимические агенты, имеющие значение в организации цикла «сна и бодрствования».
Фаза медленного сна: ГАМК, серотонин
Фаза медленного сна: норадреналин, ацетилхолин, глутамат
Бодрствование: норадреналин, глутамат, ацетилхолин, гистамин, серотонин
Новые нейрохимические агенты, имеющие значение в организации цикла «сна и бодрствования».
орексин/гипокретин
мелатонин
дельта-сон-индуцирующий пептид
аденозин
интерлейкины, мурамилпептид, цитокины
простагландины (PGD2)
Более-менее разобравшись с медленным сном, восстанавливающим силы организма, ученые бьются над загадкой сна быстрого. Самая правдоподобная гипотеза — благодаря быстрому сну мы адаптируемся к потокам новой информации.
Механизмы сна
Прямое изучение нейронов, вовлеченных в регуляцию смены сна и бодрствования, проведенное во второй половине XX века, показало, что нормальное функционирование коры мозга, обеспечивающее весь спектр сознательной деятельности человека, возможно при условии, что большинство слагающих ее нейронов находится в так называемом состоянии тонической деполяризации, когда их мембрана деполяризована на 5–10 мВ по сравнению с потенциалом покоя (–65...–70 мВ). Только в таком состоянии эти нейроны способны обрабатывать и отвечать на сигналы, приходящие к ним от других нервных клеток. Поддержание же такого состояния, в свою очередь, возможно при наличии мощныхтонических (т.е. длительных, устойчивых) воздействий со стороны определенных подкорковых структур, называемых активирующими.
Таких структур (их можно условно назвать «центрами бодрствования»), как сейчас ясно, несколько – вероятно, шесть или семь, и локализуются они на всех уровнях мозговой оси: в ретикулярной формации ствола, в области синего пятна и дорзальных ядер шва, в заднем гипоталамусе и базальных ядрах переднего мозга (рис. 1). В качестве молекул-передатчиков нейроны этих отделов мозга выделяют глутаминовую и аспарагиновую кислоты (глутамат, аспартат), ацетилхолин, норадреналин, дофамин, серотонин и гистамин.
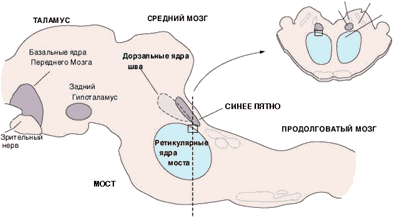
Рис. 1. Активирующие структуры мозга на схемах продольного (в центре) и поперечного (справа вверху) срезов ствола мозга кошки. Прямоугольником отмечен «центр парадоксального сна»
У человека нарушение деятельности любого из этих «центров бодрствования» не может быть скомпенсировано за счет других, несовместимо с сознанием и приводит к коме.
Казалось логичным предположить, что если в мозге есть «центры бодрствования», то должен быть и «центр сна». Однако детальное изучение свойств нейронов показало, что дело обстоит по-другому.
В разных отделах мозга разбросаны так называемые тормозные нейроны, общим для которых является выделение одного и того же химического посредника – гамма-аминомасляной кислоты, главного тормозного вещества мозга. Все эти нейроны представляют собой единый тормозной механизм, «встроенный» в систему «центров бодрствования». Механизм этот действует по принципу положительной обратной связи: стоит только активирующим нейронам по каким-то причинам ослабить свою активность, как тормозные нейроны «включаются» и ослабляют ее еще более. Так развивается процесс постепенного «углубления» сна.
Эксперименты на лабораторных животных показали, что если в бодрствовании, в состоянии тонической деполяризации, характер разрядов отдельных клеток коры мозга высоко индивидуализирован, то по мере углубления сна он коренным образом изменяется – близлежащие нейроны начинают «хором» разряжаться. При этом условия для переработки информации в мозге резко ухудшаются. Эти измерения хорошо отражают известные данные об отключении сенсорных систем и постепенном торможении психической активности по мере углубления сна у человека.
Если состояние нервных клеток во время бодрствования можно описать как состояние тонической деполяризации, то обычный, или ортодоксальный, сон (а в данном случае речь идет именно о нем) является состоянием «тонической гиперполяризации» – направление перемещения основных ионных потоков, формирующих потенциал мембраны нейрона и участвующих в проведении нервного импульса (катионов натрия, калия, кальция, анионов хлора), а также важнейших макромолекул меняется в этом случае на противоположное. Таким образом, в состоянии сна происходит восстановление мозгового гомеостаза, нарушенного в ходе предшествующего многочасового бодрствования.
Это означает подтверждение старого, как мир, но почему-то забытого правила: без хорошего сна не может быть хорошего бодрствования. Периоды тонической деполяризации и гиперполяризации должны периодически сменять друг друга для сохранения постоянства внутренней среды головного мозга и обеспечения нормального функционирования коры – субстрата высших психических функций человека.
Процесс постепенного «углубления» наступившего обычного сна развивается до определенного предела – пока не срабатывает некий «переключатель» и вся система не перебрасывается в другое состояние. Таким состоянием может быть либо пробуждение, либо наступление фазы парадоксального сна.
У большинства животных периоды сна и бодрствования на протяжении суток сменяют друг друга несколько раз, и в этом случае наступление фазы парадоксального сна обычно предшествует пробуждению. У человека периоды обычного и парадоксального сна в норме несколько раз подряд сменяют друг друга без наступления пробуждения.
Первое описание этого состояния – состояния «сна с быстрыми движениями глаз» – было сделано почти полвека назад американским физиологом Н.Клейтманом и его аспирантами Ю.Азеринским и В.Дементом. Позже огромный вклад в изучение этого явления внес Мишель Жуве. Ему же принадлежит сама формулировка понятия «парадоксальный сон».
В отличие от обычного сна сон парадоксальный имеет ярко выраженную активную природу. В этом состоянии в мозг не поступает информация от «входов» (органов чувств) и не подается информация на «выходы» (мышечную систему), однако клетки мозга проявляют чрезвычайную активность.
Получается не сон и не бодрствование, а, согласно Жуве, особое – третье – состояние организма, характеризующееся действительно парадоксальным сочетанием признаков внешнего покоя с чрезвычайно высокой активацией мозга и переживанием сновидений – как бы активное бодрствование, направленное внутрь. Открытие парадоксального сна и описание связанных с ним явлений совершило настоящую революцию в науке и изменило все представления человечества о природе сна.
Исследования позволили предположить, что в этом состоянии интенсивно перерабатывается та информация, которая была получена и сохранена в памяти в период предшествующего бодрствования. Кроме того, согласно гипотезе М.Жуве, во время парадоксального сна каким-то образом происходит переброска в нейрологическую память элементов наследственно закрепленных (инстинктивных) схем поведения. Подтверждением этому служат эмоционально окрашенные сновидения у человека, а также обнаруженный М.Жуве со своими сотрудниками и детально исследованный американским ученым Э.Моррисоном феномен демонстрации переживаемых сновидений кошками – после разрушения одной крошечной точки в задней части мозга, снимающей двигательное торможение в состоянии парадоксального сна, можно по характеру движений вполне представить себе, что именно кошка «видит» и что она «делает» во сне (охотится за мышью, убегает от собаки, дерется с соперниками и т.п.).
Запускается парадоксальный сон, как теперь установлено, из четко очерченного центра, расположенного в задней части мозга, в области варолиева моста и продолговатого мозга. Химическими передатчиками сигналов расположенных в этом «центре парадоксального сна» клеток служат ацетилхолин и глутаминовая кислота.
В 1980-е годы было обнаружено, что в парадоксальном сне те нейроны активирующих систем мозга, которые выделяют передатчики ацетилхолин и глутамат (они расположены в ретикулярной формации ствола и базальных ядрах переднего мозга), чрезвычайно активны, а нейроны, выделяющие в качестве химических передатчиков моноамины (норадреналин, серотонин и гистамин) выключаются и «молчат».
Этот факт определяет физиологическое различие между бодрствованием и парадоксальным сном, а на психическом уровне – различие между нашим восприятием внешнего мира и восприятием мира воображаемого, мира сновидений (рис. 2).
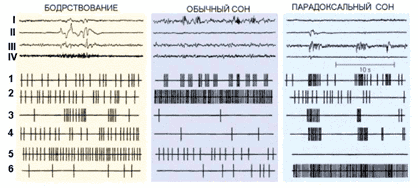
Рис. 2. Суммарная электрическая активность (I–IV) и активность одиночных нейронов (1–6) в цикле сон–бодрствование у кошки: I – кора больших полушарий, II – глаза, III – таламус, IV – мышцы шеи; 1– нейроны коры и таламуса, 2 – нейроны преоптической области переднего гипоталамуса, выделяющие гамма-аминомасляную кислоту, 3 – нейроны ретикулярных ядер моста, 4 – нейроны зрительной системы, 5 – активирующие нейроны, выделяющие моноамины (норадреналин, серотонин, гистамин), 6 – нейроны, расположенные в «центре парадоксального сна»
Регуляторы сна
Таким образом, об исполнительных механизмах трех функциональных состояний мозга – бодрствования, обычного сна и парадоксального сна – известно уже довольно много. Практически неизвестным, однако, остается биохимический механизм, управляющий чередованием этих состояний. На эту роль требуются вещества, обладающие более длительным сроком жизни в организме, чем «классические» нейропередатчики, менее специфичные по отношению к конкретным белкам-рецепторам, способные переноситься током ликвора (спинномозговой жидкости) и распространяться по межклеточной жидкости, оказывая воздействия на обширные области в мозге, иногда довольно удаленные от места выброса. Такие вещества стали известны сравнительно недавно – это регуляторные пептиды, продукты прицельного расщепления белков, эволюционно древние передатчики, широко распространенные в мозге и организме и играющие важную роль в целом ряде физиологических процессов.
Из них в первую очередь обращает на себя внимание так называемый «Дельта-Сон индуцирующий пептид» (ДСИП) – единственное вещество, выделенное из крови кроликов группой швейцарских авторов еще в конце 70-х годов как регулятор сна. В исследованиях введение этого вещества подопытным животным не давало соответствующего эффекта – возможно, в силу быстрого распада его молекул под воздействием ферментов организма. Однако ученым удалось обнаружить «снотворные» свойства у некоторых производных ДСИП – так называемых «структурных аналогов», которым химическим путем была придана повышенная устойчивость к разрушающим ферментам. Такие вещества представляют определенный интерес для дальнейшего фармакологического изучения, тем более что есть основания предполагать у них и способность усиливать сопротивляемость организма стрессорным воздействиям.
Интересно, однако, что некоторые особенности ДСИП вызывают сомнение в его природном происхождении. Этот пептид не входит ни в одно из многочисленных ныне известных пептидных семейств. Не удалось обнаружить ни его рецептора, ни гена, кодирующего белок-предшественник, из которого выщепляется ДСИП, ни самого этого белка. Возникает предположение: не является ли этот пептид своего рода артефактом, результатом ошибки в установлении аминокислотной последовательности молекулы при его выделении в ничтожных количествах (0,3 мг)? Проверить эту гипотезу не представлялось возможным вплоть до 1998 года, когда с помощью пептидного банка EROP-MOSCOW, созданного доктором биологических наук А.А. Замятниным, сотрудником Института биохимии РАН, удалось случайно обнаружить структурное сходство ДСИП и одного из дерморфинов – природных пептидов, впервые выделенных в начале 80-х годов из кожи (дермы) лягушек и обладающих сильнейшими опиоидными (морфиноподобными) свойствами.
Тогда и были разработаны структурные последовательности молекул двух таких пептидов (аналогов и ДСИП, и дерморфина одновременно) – совершенно идентичных, но являющихся оптическими изомерами. Затем эти пептиды были синтезированы в НИИ химии при Санкт-Петербургском университете. При введении в мозг подопытным кроликам один из этих пептидов оказался абсолютно неактивным, зато другой, обладающий повышенной устойчивостью по отношению к разрушающему действию ферментов мозга, продемонстрировал сильнейший эффект, заключавшийся в значительном увеличении и длительности обычного сна – как за счет уменьшения времени бодрствования, так и за счет сокращения периода сна парадоксального.
Характер этого «снотворного» эффекта очень напоминал эффект активных аналогов ДСИП, но в данном случае действие было гораздо более сильным.
Интересно, что даже этот, «устойчивый» к воздействию ферментов пептид разрушается в организме животного очень быстро – его концентрация уменьшается вдвое в течение нескольких минут. А вот эффект от его введения проявляется только через 5 часов и продолжается в течение еще 5 часов (рис. 3). Таким образом очевидно, что это вещество действует на клетки мозга не непосредственно, а запускает длинную цепь пока не известных событий. Конечным звеном этой цепи и является изменение баланса нейропередатчиков, приводящее к уменьшению времени бодрствования и увеличению времени обычного сна.
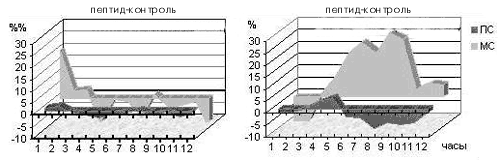
Рис. 3. Действие неактивного (слева) и активного (справа) оптических изомеров аналога дерморфина на обычный (медленноволновый, МС) и парадоксальный сон (ПС). По оси абсцисс – время от момента введения в часах. По оси ординат – разность (в %%) в продолжительности сна между опытом и контролем в среднем по группе
Открытие гипногенных (снотворных) свойств дерморфинов создает перспективы не только нового пути исследования механизмов регуляции сна, но и для создания принципиально новых лекарственных средств снотворного действия. Ведь пептиды этой группы изучены относительно неплохо – для них известны и воспринимающие рецепторы, и белки-предшественники, и гены, их кодирующие.
Кроме пептидов, чрезвычайный интерес представляет еще одна система мозга, играющая важнейшую роль в регуляции биоритмов и чередовании сна и бодрствования. Это эпифиз и выделяемый им гормон мелатонин. Эпифиз (верхний придаток мозга), являющийся «третьим глазом» у холоднокровных позвоночных и птиц, у млекопитающих утерял как способность непосредственно реагировать на свет, так и прямые нервные связи с остальным мозгом, и превратился в железу внутренней секреции. У человека эпифиз особенно активен в раннем возрасте, когда его основной функцией является, по-видимому, торможение гормонов передней доли гипофиза. В более зрелом возрасте на первый план выходит другая особенность эпифиза – его способность синтезировать мелатонин и «выбрасывать» его в строгом соответствии с внешней освещенностью, а именно в темное время суток. Информация об интенсивности окружающего освещения от сетчатки глаз поступает через зрительный нерв к супрахиазматическим ядрам преоптической области переднего гипоталамуса – «биологическим часам» организма, оттуда в «вегетативный центр» мозга, расположенный в медиальном гипоталамусе, затем через ствол и продолговатый мозг в спинной мозг и, наконец, через симпатические нервы обратно в головной мозг к пинеалоцитам – клеткам эпифиза. Интересно, что супрахиазматические ядра, в свою очередь, весьма богаты рецепторами мелатонина, т.е. обе эти структуры мозга явно взаимодействуют между собой (рис. 4).
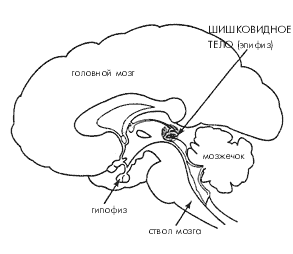
Рис. 4. Поперечный срез мозга человека (схема)
Функция эпифиза как железы внутренней секреции, выделяющей мелатонин, была установлена еще в конце 50-х годов XX века, но стала интенсивно изучаться лишь в последнее время – в связи с обширным применением синтетического мелатонина в медицине и соответствующей рекламной шумихой. Предполагается, что функция эта состоит (по крайней мере у человека) в том, чтобы обеспечить «привязку» покоя и сна к темному, а активности и бодрствования – к светлому периоду суток. И хотя исследования на людях и эксперименты на обезьянах и других млекопитающих, активных в дневное время, подтвердили прямое участие мелатонина в регуляции сна, эта гипотеза представляется все же несколько странной. Ведь синтез и выделение мелатонина происходят в темноте и блокируются на свету не только у дневных животных, но и у тех, которым свойственна ночная или сумеречная активность (рис. 5).
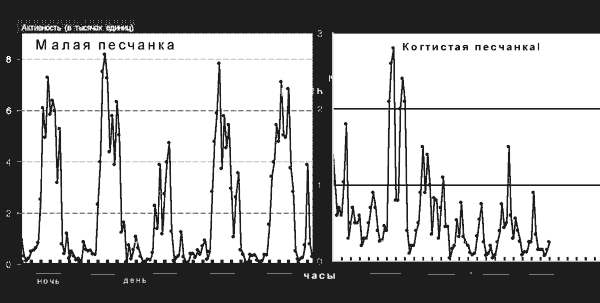
Рис. 5. Суточные ритмы активности и покоя у двух родственных видов грызунов; слева – малая песчанка, которой присуща ночная активность; справа – когтистая песчанка, активная в основном в сумерки
Экспериментами ученые провели изучение воздействия малых доз мелатонина (такие дозы обладают мягким снотворным эффектом у людей) на внутрисуточный ритм смены сна и бодрствования у подопытных кроликов, содержавшихся в условиях строгого чередования равных (12-часовых) светлых и темных периодов. Делая животным инъекцию мелатонина через час после включения света, когда уровень эндогенного («родного») гормона в их крови падает не менее чем в 15 раз по сравнению с ночным уровнем, мы ожидали обнаружить в первую очередь подавление парадоксального сна, поскольку ночью у кроликов мелатонина значительно меньше, чем днем: «много мелатонина – мало парадоксального сна», и наоборот. Однако вместо этого произошло резкое (2–3-кратное) увеличение доли парадоксального сна в течение всего 12-часового «дневного» периода (рис. 6)!
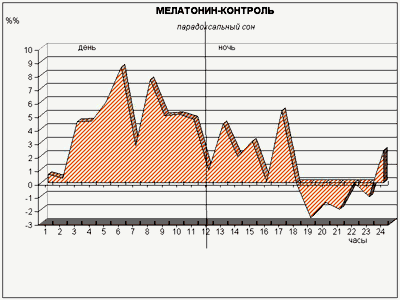
Рис. 6. Действие малой дозы мелатонина, введенной «в противофазе» с его естественной продукцией, на парадоксальный сон кроликов. Обозначения – как на рис. 3
У кроликов в лаборатории циклы сон (обычный сон, сменяемый сном пародоксальным) – бодрствование сменяют друга на протяжении примерно каждых 20 мин, вне зависимости от времени суток и освещенности. А вот соотношение обычного и пародоксального сна при этом изменяется.
За счет чего это происходит, ученым пока не ясно. Поскольку искусственно введенный в организм мелатонин, так же как и пептиды, быстро разрушается, очевидно, что и в этом случае эффект носит не «фармакологический», а «физиологический» характер. Т.е. действие оказывает не само вводимое вещество, а механизм, который это вещество запускает.
Про мелатонин известно, что он может оказывать тормозящее воздействие на определенные нейроны супрахиазматического ядра, оказывающие, в свою очередь, тормозящее влияние на «центр парадоксального сна» в задней части мозга, о котором рассказывалось ранее. Такое «торможение торможения» и может приводить к повышеннию активности этого центра. Пока это всего лишь гипотеза, но если она подтвердится, придется пересмотреть и представления о роли мелатонина в регуляции сна у человека.
Профессор Владимир Ковальзон говорит, что, если бы у животных была возможность спать целый день, они бы спали. Многие дневные животные предпочитает спать пару раз днем и всю ночь. Но ночной сон у них дробный, и после каждого цикла они просыпаются. Как правило, такой цикл у животных, как и у людей, состоит из двух фаз — медленного сна и парадоксального. У лошади, например, парадоксальный сон, по-видимому, сильно сдвинут к утру. Всю ночь она спит стоя, а под утро зачастую ложится, кладет голову на круп и видит сны.
Продолжительность цикла у различных животных разная, у некоторых она может составлять всего несколько минут. Общая же продолжительность сна животных может варьировать от двух часов у жирафа до двадцати у мешотчатого прыгуна. Макаки и шимпанзе спят около десяти часов в сутки. Коровы, лошади, кролики спят с открытыми глазами. У копытных часто наблюдается так называемый коллективный сон — пока одни особи спят, другие бодрствуют, охраняя стадо, потом ме