


Нейрон и исследование его активности
Федеральное агентство по образованию
ГОУ ВПО «САРАТОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ ИМЕНИ Н.Г.ЧЕРНЫШЕВСКОГО»
Кафедра динамического моделирования и биомедицинской инженерии
КУРСОВАЯ РАБОТА
Нейрон и исследование его активности
студент 1 курса
факультета нано- и биомедицинских технологий
Агапова Мария Геннадьевна
Научный руководитель
Профессор,д.ф.-м.н. Б.П. Безручко
Зав. кафедрой
профессор, д.ф.-м.н. Б.П. Безручко
Саратов 2006 г.
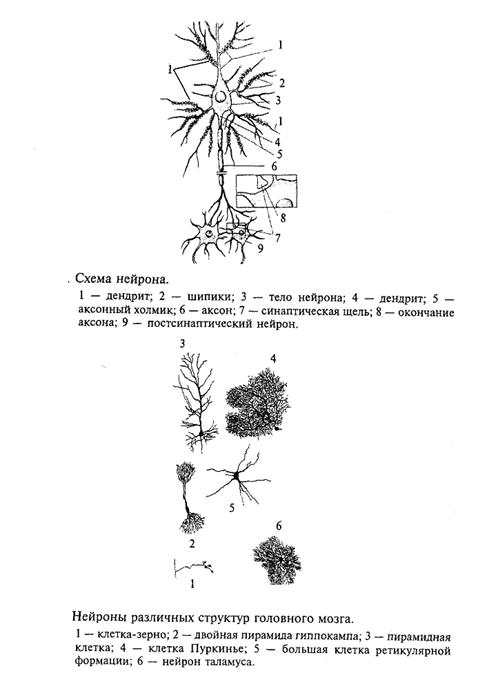
Нейрон является главной клеткой центральной нервной системы. Формы нейронов чрезвычайно многообразны, но основные части неизменны у всех типов нейронов. Нейрон состоит из следующих частей: сомы (тела) и многочисленных разветвленных отростков. У каждого нейрона есть два типа отростков: аксон, по которому возбуждение передается от нейрона к другому нейрону, и многочисленные дендриты (от греч. дерево), на которых заканчиваются синапсами (от греч. контакт) аксоны от других нейронов. Нейрон проводит возбуждение только от дендрита к аксону.
Основным свойством нейрона является способность возбуждаться (генерировать электрический импульс) и передавать (проводить) это возбуждение к другим нейронам, мышечным, железистым и другим клеткам.
На рис. 1 показана схема нейрона, на которой легко прослеживаются его основные части.
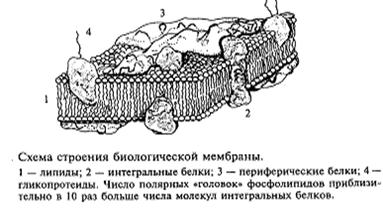
Нейроны разных отделов мозга выполняют очень разнообразную работу, и в соответствии с этим форма нейронов из разных частей головного мозга также многообразна (рис. 2). Нейроны, расположенные на выходе нейронной сети какой-то структуры, имеют длинный аксон, по которому возбуждение покидает данную мозговую структуру. Например, нейроны двигательной коры головного мозга, так называемые пирамиды Беца (названные в честь киевского анатома Б. Беца, впервые их описавшего в середине XIX века), имеют у человека аксон около 1 м, он соединяет двигательную кору больших полушарий с сегментами спинного мозга. По этому аксону передаются «двигательные команды», например «пошевелить пальцами ноги». Как возбуждается нейрон? Основная роль в этом процессе принадлежит мембране, которая отделяет цитоплазму клетки от окружающей среды. Мембрана нейрона, как и любой другой клетки, устроена очень сложно. В своей основе все известные биологические мембраны имеют однообразное строение (рис. 3): слой молекул белка, затем слой молекул липидов и еще один слой молекул белка. Вся эта конструкция напоминает два бутерброда, сложенных маслом друг к другу. Толщина такой мембраны составляет 7 - 11 нм. Чтобы представить эти размеры, вообразите, что толщина вашего волоса уменьшилась в 10 тыс. раз. В такую мембрану встроены разнообразные частицы. Одни из них являются частицами белка и пронизывают мембрану насквозь (интегральные белки), они образуют места прохождения для ряда ионов: натрия, калия, кальция, хлора. Это так называемые ионные каналы. Другие частицы прикреплены на внешней поверхности мембраны и состоят не только из молекул белка, но и из полисахаридов. Это рецепторы для молекул биологически активных веществ, например медиаторов, гормонов и др. Часто в состав рецептора, кроме места для связывания специфической молекулы, входит и ионный канал.
Главную роль в возбуждении нейрона играют ионные каналы мембраны. Эти каналы бывают двух видов: одни работают постоянно и откачивают из нейрона ионы натрия и накачивают в цитоплазму ионы калия. Благодаря работе этих каналов (их называют еще насосными каналами или ионным насосом), постоянно потребляющих энергию, в клетке создается разность концентраций ионов: внутри клетки концентрация ионов калия примерно в 30 раз превышает их концентрацию вне клетки, тогда как концентрация ионов натрия в клетке очень небольшая - примерно в 50 раз меньше, чем снаружи клетки. Свойство мембраны постоянно поддерживать разность ионных концентраций между цитоплазмой и окружающей средой характерно не только для нервной, но и для любой клетки организма. В результате между цитоплазмой и внешней средой на мембране клетки возникает потенциал: цитоплазма клетки заряжается отрицательно на величину около 70мВ относительно внешней среды клетки. Измерить этот потенциал можно в лаборатории стеклянным электродом, если в клетку ввести очень тонкую (меньше 1 мкм) стеклянную трубочку, заполненную раствором соли. Стекло в таком электроде играет роль хорошего изолятора, а раствор соли - проводника. Электрод соединяют с усилителем электрических сигналов и на экране осциллографа регистрируют этот потенциал. Оказывается, потенциал порядка - 70 мВ сохраняется в отсутствие ионов натрия, но зависит от концентрации ионов калия. Другими словами, в создании этого потенциала участвуют только ионы калия, в связи, с чем этот потенциал получил название «калиевый потенциал покоя», или просто «потенциал покоя». Таким образом, это потенциал любой покоящейся клетки нашего организма, в том числе и нейрона.
ВОЗБУЖДЕНИЕ НЕЙРОНА
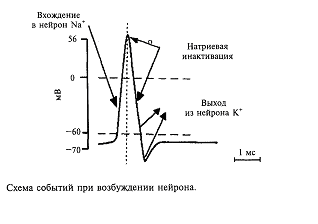
Нейрон в отличие от других клеток способен возбуждаться. Под возбуждением нейрона понимают генерацию нейроном потенциала действия. Основная роль в возбуждении принадлежит другому типу ионных каналов, при открытии которых ионы натрия устремляются в клетку. Напомним, что благодаря постоянной работе насосных каналов концентрация натриевых ионов вне клетки примерно в 50 раз больше, чем в клетке, поэтому при открытии натриевых каналов ионы натрия устремляются в клетку, а ионы калия через открытые калиевые каналы начинают выходить из клетки. Для каждого типа ионов - натрия и калия - имеется свой собственный тип ионного канала. Движение ионов по этим каналам происходит по концентрационным градиентам, т.е. из места высокой концентрации в место с более низкой концентрацией.
Ответим на вопрос: как ионные каналы открываются и закрываются? В покоящемся нейроне натриевые каналы мембраны закрыты и на мембране, как это уже описывалось выше, регистрируется потенциал покоя порядка-70 мВ (отрицательность в цитоплазме). Если потенциал мембраны деполяризовать (уменьшить поляризацию мембраны) примерно на 10 мВ, натриевый ионный канал открывается (рис. 4). Действительно, в канале имеется своеобразная заслонка, которая реагирует на потенциал мембраны, открывая этот канал при достижении потенциала определенной величины. Такой канал называется потенциалзависимым. Как только канал открывается, в цитоплазму нейрона устремляются из межклеточной среды ионы натрия, которых там примерно в 50 раз больше, чем в цитоплазме. Такое движение ионов является следствием простого физического закона: ионы движутся по концентрационному градиенту. Таким образом, в нейрон поступают ионы натрия, они заряжены положительно. Другими словами, через мембрану будет протекать входящий ток ионов натрия, который будет смещать потенциал мембраны в сторону деполяризации, т. е. уменьшать поляризацию мембраны. Чем больше ионов натрия войдет в цитоплазму нейрона, тем больше его мембрана деполяризуется. Потенциал на мембране будет увеличиваться, открывая все большее количество натриевых каналов. Но этот потенциал будет расти не бесконечно, а только до тех пор, пока не станет равным примерно +55 мВ. Этот потенциал соответствует присутствующим в нейроне и вне его концентрациям ионов натрия, поэтому его называют натриевым равновесным потенциалом. Вспомним, что в покое мембрана имела потенциал -70 мВ, тогда абсолютная амплитуда потенциала составит величину около 125 мВ. Мы говорим «около», «примерно» потому, что у клеток разного размера и типов этот потенциал может несколько отличаться, что связано с формой этих клеток (например, количеством отростков), а также с особенностями их мембран.
Все изложенное выше можно формально описать следующим образом. В покое клетка ведет себя как «калиевый электрод», а при возбуждении - как «натриевый электрод». Однако после того как потенциал на мембране достигнет своего максимального значения +55 мВ, натриевый ионный канал со стороны, обращенной в цитоплазму, закупоривается специальной белковой молекулой. Это так называемая «натриевая инактивация» (см. рис.4); она наступает примерно через 0,5-1 мс и не зависит от потенциала на мембране. Мембрана становится непроницаемой для натриевых ионов. Для того чтобы потенциал мембраны вернулся к исходному состоянию-состоянию покоя, необходимо, чтобы из клетки выходил ток положительных частиц. Такими частицами в нейронах являются ионы калия. Они начинают выходить через открытые калиевые каналы. Вспомните, что в клетке в состоянии покоя накапливаются ионы калия, поэтому при открывании калиевых каналов эти ионы покидают нейрон, возвращая мембранный потенциал к исходному уровню (уровню покоя). В результате этих процессов мембрана нейрона возвращается к состоянию покоя (-70 мВ) и нейрон готовится к следующему акту возбуждения.
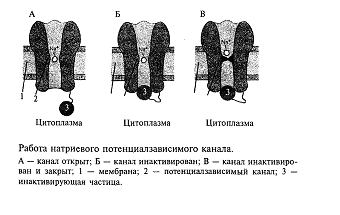
Таким образом, выражением возбуждения нейрона является генерация на мембране нейрона потенциала действия. Его длительность в нервных клетках составляет величину около 1/1000с(1 мс). Описанная последовательность событий приведена на рис. 5.
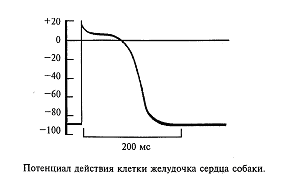
Подобные потенциалы действия могут возникать и в других клетках, назначение которых - возбуждаться и передавать это возбуждение другим клеткам. Например, сердечная мышца имеет в своем составе специальные мышечные волокна, обеспечивающие бесперебойную работу сердца в автоматическом режиме. В этих клетках также генерируются потенциалы действия (рис. 6). Однако они имеют затянутую, почти плоскую вершину, и длительность такого потенциала действия может затянуться до нескольких сот миллисекунд (сравните с 1 мс у нейрона). Такой характер потенциала действия мышечной клетки сердца физиологически оправдан, так как возбуждение сердечной мышцы должно быть длительным, чтобы кровь успела покинуть желудочек. С чем же связан такой затянутый потенциал действия у этого типа клетки? Оказалось, в мембране этих клеток натриевые ионные каналы не так быстро закрываются, как в нейронах, т. е. натриевая инактивация затянута.
Как ясно из этого описания, возбуждение (потенциал действия) нейрона сменяется так называемым «покоем». Однако никакого покоя в этот период нет. Как уже указывалось выше, в мембране есть еще и насосные каналы, количество которых примерно в 10 раз больше ионных, и они постоянно работают, откачивая из цитоплазмы излишек ионов натрия и закачивая туда недостающие ионы калия. Благодаря неустанной работе этих каналов нейрон всегда готов к возбуждению.
Описанный выше механизм возбуждения клетки (конечно, далеко не все клетки нашего организма способны возбуждаться) в основных чертах одинаков не только в нейронах и мышечных клетках человека, но и в аналогичных клетках других организмов. Например, в нейронах моллюсков, червей, крыс и обезьян при возбуждении происходят описанные выше последовательности событий. Более того, конструкция мембран, включая каналы, также примерно одинакова у всех организмов Земли.
Как уже указывалось, каналы представляют собой белковые молекулы, «прошивающие» мембрану (одна часть молекулы находится в цитоплазме, а другая во внеклеточной среде). Интересно, что эти белковые молекулы, образующие ионный или насосный каналы, не вечны, а постоянно заменяются на новые (примерно каждые несколько часов). Все это свидетельствует об очень большой динамичности структуры нейрона.
Нейрон способен к возбуждению, которое состоит в том, что мембрана нейрона в состоянии покоя имеет потенциал порядка - 70 мВ (отрицательность в цитоплазме), а в состоянии возбуждения приобретает потенциал +55 мВ. Таким образом, абсолютная величина потенциала действия - около 125 мВ. Длительность потенциала действия нейрона составляет всего около 1 мс (1/1000 с).
Далее это возбуждение (потенциал действия) должно передаться другому нейрону или какой-то другой клетке, например мышечной, железистой и др.
ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ
Возбуждение в виде потенциала действия покидает тело нейрона по его отростку, который называется аксоном. Аксоны отдельных нейронов обычно объединяются в пучки - нервы, а сами аксоны в этих пучках называются нервными волокнами. Природа позаботилась, чтобы волокна максимально хорошо справлялись с функцией проведения возбуждения в виде потенциалов действия. Для этой цели отдельные нервные волокна (аксоны отдельных нейронов) имеют специальные чехлы, выполненные из хорошего электрического изолятора (см. рис. 1). Чехол прерывается примерно через каждые 0,5-1,5 мм; это связано с тем, что отдельные участки чехла образуются в результате того, что специальные клетки в очень ранний период развития организма (в основном еще до рождения) обволакивают небольшие участки аксона. На рис.7 показано, как это происходит. В периферических нервах миелин образуется клетками, которые получили название шванновских, а в головном мозге это происходит за счет клеток олигодендроглии.
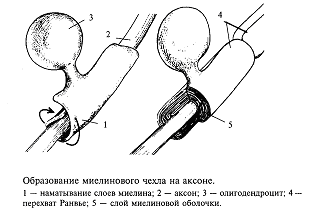
Этот процесс называется миелинизацией, так как в результате образуется чехол из вещества миелина, примерно на 2/3 состоящего из жира и являющегося хорошим электрическим изолятором. Исследователи придают очень большое значение процессу миелинизации в развитии мозга.
Известно, что у новорожденного ребенка миелинизировано примерно 2/3 волокон головного мозга. Примерно к 12 годам завершается следующий этап миелинизации. Это соответствует тому, что у ребенка уже формируется функция внимания, он достаточно хорошо владеет собой. Вместе с тем полностью процесс миелинизации заканчивается только при завершении полового созревания. Таким образом, процесс миелинизации является показателем созревания ряда психических функций. В то же время известны заболевания нервной системы человека, которые связаны с демиелинизацией нервных волокон, что сопровождается тяжелыми страданиями. К самым известным относится рассеянный склероз. Это заболевание развивается незаметно и очень медленно, последствием является паралич движения.
Почему же так важна миелинизация нервных волокон? Оказывается, миелинизированные волокна в сотни, раз быстрее проводят возбуждение, чем немиелинизированные, т. е. нейронные сети нашего мозга могут работать с большей скоростью, а значит, более эффективно. Поэтому не миелинизируются в нашем организме только самые тонкие волокна (менее 1 мкм в диаметре), которые проводят возбуждение к медленно работающим органам кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре.
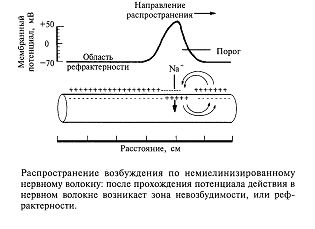
Как происходит распространение возбуждения по нервному волокну? Вначале разберем случай немиелинизированного нервного волокна. На рис.8 показана схема нервного волокна. Возбужденный участок аксона характеризуется тем, что мембрана, обращенная к аксоплазме, заряжается положительно относительно экстраклеточной среды. Невозбужденные (покоящиеся) участки мембраны волокна отрицательны внутри. Между возбужденным и невозбужденным участками мембраны возникает разность потенциалов и начинает протекать ток. На рисунке это отражено линиями тока, пересекающими мембрану со стороны аксоплазмы,-выходящий ток, который деполяризует соседний невозбужденный участок волокна. Возбуждение движется по волокну только в одном направлении (показано стрелкой) и не может пойти в другую сторону, так как после возбуждения участка волокна в нем наступает рефрактерность - зона невозбудимости. Нам уже известно, что деполяризация приводит к открыванию потенциалзависимых натриевых каналов и в соседнем участке мембраны развивается потенциал действия. Затем натриевый канал инактивируется и закрывается, что и приводит к зоне невозбудимости волокна. Эта последовательность событий повторяется для каждого соседнего участка волокна. На каждое такое возбуждение тратится определенное время. Специальные исследования показали, что скорость проведения возбуждения немиелинизированных волокон пропорциональна их диаметру: чем больше диаметр, тем выше скорость движения импульсов. Например, немиелинизированные волокна, проводящие возбуждение со скоростью 100 - 120 м/с, должны иметь диаметр около 1000 мкм (1 мм).
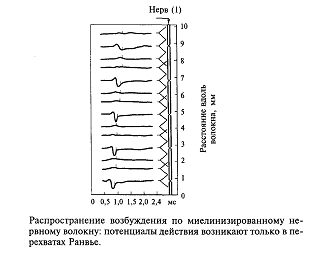
У млекопитающих животных природа сохранила немиелинизированными только те возбуждение о боли, температуре, управляют медленно работающими внутренними органами мочевым волокна, которые проводят органами - мочевым пузырем, кишечником и пр. Практически все нервные волокна в центральной нервной системе человека имеют миелиновые чехлы. На рис.9 показано, что если вдоль волокна, покрытого миелином, регистрировать прохождение возбуждения, то потенциал действия возникает только в перехватах Ранвье. Оказывается, миелин, являясь хорошим электрическим изолятором, не пропускает выхода линий тока от предшествующего возбужденного участка. Выход тока в этом случае возможен только через те участки мембраны, которые находятся на стыке между двумя участками миелина. Напомним, что каждый участок образован только одной клеткой, поэтому это стыки между двумя клетками, образующими соседние участки миелиновой оболочки. Мембрана аксона между двумя соседними миелиновыми чехлами оказывается не покрытой миелином (так называемый перехват Ранвье). Благодаря такому устройству мембрана волокна возбуждается только в местах перехватов Ранвье. Вследствие этого потенциал действия (возбуждение) как бы перескакивает через участки изолированной мембраны. Другими словами, возбуждение движется скачками от перехвата к перехвату. Это похоже на те волшебные сапоги-скороходы, которые надевал кот в известной сказке, мгновенно переносясь из одного места в другое.
СИНАПС
Как передается возбуждение от одного нейрона другому или от нейрона, например, на мышечное волокно? Этой проблемой интересуются не только профессиональные нейробиологи, но и врачи, особенно фармакологи. Знание биологических механизмов необходимо для лечения некоторых заболеваний, а также для создания новых лекарств и препаратов. Дело в том, что одними из основных мест воздействия этих веществ на организм человека являются места передачи возбуждения с одного нейрона на другой (или на другую клетку, например клетку сердечной мышцы, стенки сосудов и пр.). Отросток нейрона аксон направляется к другому нейрону и образует на нем контакт, который называют синапсом (в переводе с греческого - контакт; см. рис. 1). Именно синапс хранит многие тайны мозга. Нарушение этого контакта, например, веществами, блокирующими его работу, приводит к тяжелейшим последствиям для человека. Это место приложения действия наркотиков. Примеры будут приведены ниже, а сейчас рассмотрим, как устроен и как работает синапс.
Трудности этого исследования определяются тем, что сам синапс очень маленький (его диаметр не более 1 мкм). Один нейрон получает такие контакты, как правило, от нескольких тысяч (3 - 10 тыс.) других нейронов. Каждый синапс надежно закрыт специальными клетками глии, поэтому исследовать его очень непросто. На рис. 10 показана схема синапса, как это представляет себе современная наука. Несмотря на свою миниатюрность, он устроен весьма сложно. Одним из его основных компонентов являются пузырьки, которые находятся внутри синапса. Эти пузырьки содержат биологически очень активное вещество, которое называется нейротрансмиттером, или медиатором (передатчиком).
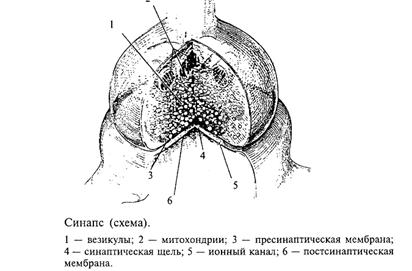
Вспомним, что нервный импульс (возбуждение) с огромной скоростью продвигается по волокну и подходит к синапсу. Этот потенциал действия вызывает деполяризацию мембраны синапса (рис. 11), однако это не приводит к генерации нового возбуждения (потенциала действия), а вызывает открывание специальных ионных каналов, с которыми мы еще не знакомы. Эти каналы пропускают ионы кальция внутрь синапса. Ионы кальция играют очень большую роль в деятельности организма. Специальная железа внутренней секреции - паращитовидная (она находится поверх щитовидной железы) регулирует содержание кальция в организме. Многие заболевания связаны с нарушением обмена кальция в организме. Например, его недостаток приводит к рахиту у маленьких детей.
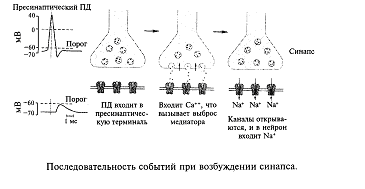
Каким образом кальций участвует в работе синапса? Попадая в цитоплазму синаптического окончания, кальций входит в связь с белками, образующими оболочку пузырьков, в которых хранится медиатор. В конечном итоге мембраны синаптических пузырьков сжимаются, выталкивая свое содержимое в синаптическую щель. Этот процесс очень напоминает сокращение мышечного волокна в мышце, во всяком случае, эти два процесса имеют одинаковый механизм на молекулярном уровне. Таким образом, связывание кальция белками оболочки пузырька приводит к ее сокращению, и содержание пузырька впрыскивается (экзоцитоз) в щель, которая отделяет мембрану одного нейрона от мембраны другого. Эта щель называется синоптической щелью. Из описания должно быть ясно, что возбуждение (электрический потенциал действия) нейрона в синапсе превращается из электрического импульса в импульс химический. Другими словами, каждое возбуждение нейрона сопровождается выбросом в окончании его аксона порции биологически активного вещества - медиатора. Далее молекулы медиатора связываются с специальными белковыми молекулами, которые находятся на мембране другого нейрона. Эти молекулы называются рецепторами. Рецепторы устроены уникально и связывают только один тип молекул. В некоторых описаниях указывается, что они подходят, как «ключ к замку» (ключ подходит только к своему замку).
Рецептор состоит из двух частей. Одну можно назвать «узнающим центром», другую - «ионным каналом». Если молекулы медиатора заняли определенные места (узнающий центр) на молекуле рецептора, то ионный канал открывается, и ионы начинают входить в клетку (ионы натрия) или выходить (ионы калия) из клетки. Другими словами, через мембрану протекает ионный ток, который вызывает изменение потенциала на мембране. Этот потенциал получил название постсинаптического потенциала (рис. 2.13). Очень важным свойством описанных ионных каналов является то, что количество открытых каналов определяется количеством связанных молекул медиатора, а не потенциалом на мембране, как в случае с электровозбудимой мембраной нервного волокна. Таким образом, постсинаптические потенциалы имеют свойство градуальности: амплитуда потенциала определяется количеством молекул медиатора, связанного рецепторами. Благодаря этой зависимости амплитуда потенциала на мембране нейрона развивается пропорционально количеству открытых каналов.
На мембране одного нейрона могут одновременно находиться два вида синапсов: тормозные и возбудительные. Все определяется устройством ионного канала мембраны. Мембрана возбудительных синапсов пропускает как ионы натрия, так и ионы калия. В этом случае мембрана нейрона деполяризуется. Мембрана тормозных синапсов пропускает только ионы хлора и гиперполяризуется. Очевидно, что если нейрон заторможен, потенциал мембраны увеличивается (гиперполяризация). Таким образом, нейрон благодаря воздействию через соответствующие синапсы может возбудиться или прекратить возбуждение, затормозиться. Все эти события происходят на соме и многочисленных отростках дендрита нейрона, на последних находится до нескольких тысяч тормозных и возбудительных синапсов.
В качестве примера разберем, как действует в синапсе медиатор, который называется ацетилхолином. Этот медиатор широко распространен в головном мозге и в периферических окончаниях нервных волокон. Например, двигательные импульсы, которые по соответствующим нервам приводят к сокращению мышц нашего тела, оперируют ацетилхолином. Ацетилхолин был открыт в 30-х годах австрийским ученым О. Леви. Эксперимент был очень прост: изолировали сердце лягушки с подходящим к нему блуждающим нервом. Было известно, что электрическая стимуляция блуждающего нерва приводит к замедлению сокращений сердца вплоть до полной его остановки. О. Леви простимулировал блуждающий нерв, получил эффект остановки сердца и взял из сердца немного крови. Оказалось, что если эту кровь добавить в желудочек работающего сердца, то оно замедляет свои сокращения. Был сделан вывод: при стимуляции блуждающего нерва выделяется вещество, останавливающее сердце. Это и был ацетилхолин. Позже был открыт фермент, который расщеплял ацетилхолин на холин (жир) и уксусную кислоту, в результате чего прекращалось действие медиатора. Этим исследованием впервые была установлена точная химическая формула медиатора и последовательность событий в типичном химическом синапсе. Эта последовательность событий сводится к следующему.
Потенциал действия, пришедший по пресинаптическому волокну к синапсу, вызывает деполяризацию, которая включает кальциевый насос, и ионы кальция поступают в синапс; ионы кальция связываются белками мембраны синаптических пузырьков, что приводит к активному опорожнению (экзоцитозу) пузырьков в синаптическую щель. Молекулы медиатора связываются (узнающим центром) соответствующими рецепторами постсинаптической мембраны, при этом открывается ионный канал. Через мембрану начинает протекать ионный ток, что приводит к возникновению на ней постсинаптического потенциала. В зависимости от характера открытых ионных каналов возникает возбудительный (открываются каналы для ионов натрия и калия) или тормозной (открываются каналы для ионов хлора) постсинаптический потенциал.
Ацетилхолин весьма широко распространен в живой природе. Например, он находится в стрекательных капсулах крапивы, в стрекательных клетках кишечнополостных животных (например, пресноводной гидры, медузы) и пр. В нашем организме ацетилхолин выбрасывается в окончаниях двигательных нервов, управляющих мышцами, из окончаний блуждающего нерва, который управляет деятельностью сердца и других внутренних органов. Человек давно знаком с антагонистом ацетилхолина - это яд кураре, которым пользовались индейцы Южной Америки при охоте на животных. Оказалось, что кураре, попадая в кровь, вызывает обездвиживание животного, и оно погибает фактически от удушья, но кураре не останавливает сердце. Исследования показали, что в организме существуют два типа рецепторов к ацетилхолину: один успешно связывает никотиновую кислоту, а другой - мускарин (вещество, которое выделено из гриба рода Muscaris). На мышцах нашего тела находятся рецепторы никотинового типа к ацетилхолину, тогда как на сердечной мышце и нейронах головного мозга - рецепторы к ацетилхолину мускаринового типа.
В настоящее время в медицине широко применяют синтетические аналоги кураре для обездвиживания больных во время сложных операций на внутренних органах. Применение этих средств приводит к полному параличу двигательной мускулатуры (связывается рецепторами никотинового типа), но не влияет на работу внутренних оранов, в том числе сердца (рецепторы мускаринового типа). Нейроны головного мозга, возбуждаемые через мускариновые ацетилхолиновые рецепторы, играют большую роль в проявлении некоторых психических функций. Сейчас известно, что гибель таких нейронов приводит к старческому слабоумию (болезнь Альцгеймера). Другим примером, который должен показать важность именно рецепторов никотинового типа на мышце к ацетилхолину, может служить заболевание, называемое miastenia grevis (мышечная слабость). Это генетически наследуемая болезнь, т. е. ее происхождение, связано с «поломками» генетического аппарата, которые передаются по наследству. Заболевание проявляется в возрасте ближе к половозрелости и начинается с мышечной слабости, которая постепенно усиливается и захватывает все более обширные группы мышц. Причиной этого недуга оказалось то, что организм больного вырабатывает белковые молекулы, которые прекрасно связываются ацетилхолиновыми рецепторами никотинового типа. Занимая эти рецепторы, они препятствуют связыванию с ними молекул ацетилхолина, выбрасываемых из синаптических окончаний двигательных нервов. Это и приводит к блокированию синаптического проведения к мышцам и, следовательно, к их параличу.
Описанный на примере ацетилхолина тип синаптической передачи - не единственный в ЦНС. Второй тип синаптической передачи также широко распространен, например, в синапсах, в которых медиаторами являются биогенные амины (дофамин, серотонин, адреналин и др.). В этом типе синапсов имеет место следующая последовательность событий. После того как образовался комплекс «молекула медиатора - рецепторный белок», активируется специальный мембранный белок (G-белок). Одна молекула медиатора при связывании с рецептором может активировать много молекул G-белка, и это усиливает эффект медиатора. Каждая активированная молекула G-белка в одних нейронах может открывать ионный канал, а в других активировать внутри клетки синтез специальных молекул, так называемых вторичных посредников. Вторичные посредники могут запускать в клетке многие биохимические реакции, связанные с синтезом, например, белка, в этом случае возникновения электрического потенциала на мембране нейрона не происходит.
Существуют и другие медиаторы. В головном мозге в качестве медиаторов «работает» целая группа веществ, которые объединены под названием биогенные амины. В середине прошлого столетия английский врач Паркинсон описал болезнь, которая проявлялась как дрожательный паралич. Это тяжелое страдание вызвано разрушением в мозге больного нейронов, которые в своих синапсах (окончаниях) выделяют дофамин - вещество из группы биогенных аминов. Тела этих нейронов находятся в среднем мозге, образуя там скопление, которое называется черной субстанцией. Исследования последних лет показали, что дофамин в мозге млекопитающих также имеет несколько типов рецепторов (в настоящее время известно шесть типов). Другое вещество из группы биогенных аминов - серотонин (другое название 5-окситриптамин) - вначале было известно как средство, приводящее к подъему кровяного давления (сосудосуживающее). Обратите внимание, что, это отражено в его названии. Однако оказалось, что истощение в головном мозге серотонина приводит к хронической бессоннице. В опытах на животных было установлено, что разрушение в мозговом стволе (задних отделах мозга) специальных ядер, которые известны в анатомии как ядра шва, приводит к хронической бессоннице и в дальнейшем гибели этих животных. Биохимическое исследование установило, что нейроны ядер шва содержат серотонин. У пациентов, страдающих хронической бессонницей, также обнаружено снижение концентрации серотонина в мозге.
К биогенным аминам относят также адреналин и норадреналин, которые содержатся в синапсах нейронов автономной нервной вегетативной системы. Во время стресса под влиянием специального гормона - адренокортикотропного (подробнее см. ниже) - из клеток коры надпочечников в кровь также выбрасываются адреналин и норадреналин.
ЭЛЕКТРОЭНЦЕФАЛОГРАФ
Необходимо специально подчеркнуть, что деятельность высшего из известных творений Эволюции — человеческого мозга неимоверно сложна и крайне трудно поддается изучению и систематизации. Поэтому, в отличие от большинства других физиологических сигналов, при анализе ЭЭГ используется намного меньше устойчивых числовых характеристик и детерминированных диагностических критериев, а адекватность клинического заключения в огромной степени определяется профессиональным опытом и внутренним чутьем высококвалифицированного клинициста.
Более того, если после анализа других электрофизиологических показателей обычно можно прописать лечение в той или иной мере восстанавливающее нормальное состояние, то результатом исследования ЭЭГ обычно являются лишь рекомендации по коррекции поведения и ограничение видов деятельности (1, стр. 127).
Коре головного мозга свойственна постоянная электрическая активность, являющаяся результатом генерации синаптических потенциалов и импульсных разрядов в отдельных нервных клетках.
Генерация в коре электрических колебаний была обнаружена Р. Катоном и А. Данилевским. Возможность регистрации биопотенциалов непосредственно от поверхности головы животных была показана В. Правдич-Неминским в 1925 г. В 1929 г. Г. Бергер зарегистрировал электрическую активность от поверхности головы человека — электроэнцефалограмму (ЭЭГ) (2, стр 147).
Электроэнцефалограмма - графическая запись биоэлектрических процессов мозга, отводимых с помощью электродов, расположенных на поверхности головы.
Ранее считали, что ЭЭГ характеризует суммарное воздействие потенциалов действия различных клеток тела и нервных волокон, расположенных вблизи измерительных электродов. Однако в настоящее время считают, что, так как пути распространения и временные соотношения между потенциалами действия в мозге почти случайны по своей, природе, то потенциалы действия вносят лишь незначительный вклад в ЭЭГ. В соответствии с принятой теорией биоэлектрические потенциалы, появляющиеся на поверхности кожи головы, характеризуют воздействие синхронизированных постсинаптическик (переходных) потенциалов (graded potentials) в различных комбинациях нейронов мозга.
Так как на окончания аксонов непрерывно поступают потенциалы действия, то потенциал покоя каждого нейрона мозга непрерывно изменяется. Эти изменения потенциалов покоя, которые происходят медленнее, чем изменения потенциалов действия, называются постсинаптическими или переходными потенциалами. Часто переходные потенциалы большого числа нейронов данной области мозга на какой-то период времени синхронизируются. Затем такая синхронизация нарушается, и возникают другие синхронизированные комплексы возбуждений, может быть, и на других частотах.
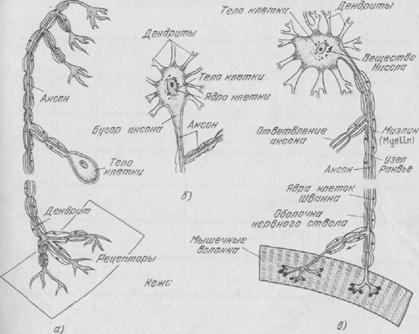
Рис. 12. Схематическое изображение трех различных типов нейронов: а) — спинальный сенсорный нейрон; б) — ассоциативный нейрон; в)— спинальный двигательный нейрон.
Как считают в настоящее время, такие синхронизированные переходные потенциалы и являются основными источниками потенциалов ЭЭГ. Типичные образцы записи ЭЭГ показаны на рис. 13. Эти потенциалы ЭЭГ, измеренные на поверхности кожи головы, в действительности представляют собой результат комбинированного (суммарного) воздействия постсинаптических (переходных) потенциалов в достаточно широкой области (коры головного мозга и в различных точках ниже ее. В общем случае эти потенциалы почти случайны по природе. Однако некоторые характеристики кривых ЭЭГ связаны с состоянием сна или патологической активностью. (4, стр. 88-92)
РИТМЫ
Под понятием "ритм" на ЭЭГ подразумевается определенный тип электрической активности, соответствующий некоторому определенному состоянию мозга и связанный с определенными церебральными механизмами. В клинических исследованиях обычно выделяют четыре типа ритмов, последовательно возрастающей частоты: дельта, тета, альфа и бета ритмы. В спокойном состоянии у человека в большей части коры больших полушарий регистрируется регулярный ритм с частотой около 8—13 Гц в секунду (альфа-ритм). В состоянии активной деятельности он сменяется частыми (более 13 в секунду) колебаниями небольшой амплитуды (бета-ритм). Во время сна он сменяется медленными (0,5—3.5 в секунду) колебаниями (дельта-ритм).
Аналогичные изменения происходят при активации восходящих путей, проецирующихся в кору. Таким образом, ЭЭГ позволяет судить о функциональном состоянии коры, например о глубине наркоза, о наличии в определенных ее зонах патологических процессов.
Для анализа деятельности корковых структур, в особенности у животных, возможно отведение потенциалов от отд