


Морфология колонии и распределительная система у двух видов герматипных кораллов рода acropora
Марфенин Н. Н.
Материал собран в 1979 г. на Большом Барьерном рифе Австралии. Показано, что в колонии у акрапор имеются, по крайней мере, четыре морфо-функциональные зоны: верхушки ветвей; сами ветви; стволы; основание колонии или зона инкрустации. Приведены характеристики каждой зоны. В пористом скелете ветви выделено пять подсистем полостей, подчиняющихся радиальной симметрии и составляющих морфологическую основу распределительной системы. Функционирование распределительной системы изучено с помощью красителей и инъекции мелких частиц угля. Внутри ветви обнаружено два продольных противоположно направленных течения: по стороне ветви, изначально (т. е. при естественном положении колонии) обращенной вверх, внутреннее течение идет дистально к верхушке; по противоположной стороне (обращенной к субстрату) проксималыю, к основанию. Колонию надо рассматривать как целостное образование; исследование морфологии целой колонии у кораллов позволит существенно обогатить их систематику новыми признаками,
Строение колонии у герматипных кораллов, в отличие от такового отдельных кораллитов, изучено очень мало. Это объясняется отчасти методическими трудностями, связанными со сложным переплетением мягкого тела и скелета, а отчасти исторически сложившимся представлением о колонии как ассоциации особей—членов колонии, в которой связи между зооидами имеют второстепенное значение, а соответственно морфология связующих структур вариабельна и не заслуживает специального рассмотрения. Поэтому в работах по морфологии кораллов можно встретить лишь описания кораллитов, а сведения о характере связей между кораллитами, строении колонии и различиях морфологии кораллитов в зависимости от места их положения в колонии практически отсутствуют. Нет данных и об анатомии распределительной системы в колонии. Систематика герматипных кораллов также основана на морфологии кораллитов, что обедняет ее возможности. Акропоры представлены в коллекциях, как правило, лишь фрагментами колоний, по которым нельзя изучить морфологию колонии в целом.
Настоящее исследование имело целью, во-первых, выделить закономерные особенности формы колонии, ее морфологически и функционально отличные части, а во-вторых, охарактеризовать связи между кораллитамн, т. е. описать распределительную систему колонии.
Сбор материала и экспериментальная работа проведены на Биологической станции на о-ве Херон, расположенном у южной оконечности Большого Барьерного Рифа Австралии, в 1979 г. при поддержке кафедры зоологии Австралийского национального университета. Работа стала возможной благодаря осуществлению соглашения между Московским университетом и Австралийским национальным университетом (Канберра) по научному обмену.
Исследование ограничивалось рядом видов рода Асгорога, представители которого играют важную роль на коралловых рифах. Подробнее других были исследованы A. fomosa (Dana, 1846), Л. aspera (Dana, 1846). На их примере и дается описание общей морфологии колонии.
Материал был собран на риф-флэте вблизи биостанции. Для содержания колоний использовали проточные аквариумы. Морфологию скелета изучали на продольных и поперечных сколах, в основном на живом материале. Ткани в некоторых случаях окрашивали витальным красителем нильский голубой. Для выявления зон роста использовали ализарин красный-С, который включается в скелет пропорционально интенсивности процесса скелетообразования (Lamberts, 1974, 1978).
Для изучения функционирования распределительной системы были применены простые методики инъекции толченого угля в живые колонии, окраски скола витальным красителем, наблюдения за выносом зооксантелл в экспериментальных условиях, описание которых дано при дальнейшем изложении.
Определение кораллов проводили по монография Воллас (Wallace, I978) и на основе ознакомления с коллекциями Квинслсндского музея (коллекция Кадеп Воллас) и Австралийского института морских наук (коллекция Дж. Всрона).
Подразделeние колонии на основные части
Акропоры (колонии из рода Асгорога) имеют ветвистое строение. Многочисленные мелкие радиальные кораллиты располагаются латерально на ветвях. Венчает каждую ветвь несколько более крупный аксиальный кораллит. Колония в общем случае отчетливо подразделена на четыре основные морфо-функциональные зоны или части: 1) верхушки ветвей; 2) сами ветви; 3) ствол (или стволы) колонии; 4) основание колонии или зона инкрустации. Каждая часть имеет свою специфику и играет определенную роль в колонии.
Верхушки ветвей. В них осуществляются продольный рост (в длину), а также наиболее интенсивный поперечный рост (утолщение ветви) и морфогенез радиальных кораллитов 1-го порядка. Удлинение ветвей происходит путем надстраивания апикальной части аксиального кораллита (рис. 1, Л, 1). В самой его верхушке находится зона наиболее интенсивного роста.
По бокам аксиального кораллита происходит надстраиваиие кост (пластинковидных радиально расположенных элементов скелета), в процессе которого образуются радиальные кораллиты 1-го порядка (рис. 1, Л, 2). Проксимальнее рост радиальных кораллитов 1-го порядка и скелета ветви продолжается на основе созданной аксиальным кораллитом матрицы. Диаметр ветви поэтому увеличивается в проксимальном направлении. Однако скорость роста в ширину в старых частях ветви уменьшается, так что в профиле любой ветви можно более или менее отчетливо выделить точку перегиба, которую можно использовать для установления границы верхушки как особой морфо-функциональной структуры.
Показатель перегиба в профиле ветви имеет практическое значение в основном при работе с коллекциями скелетов кораллов, а при работе с живым материалом можно использовать и другой критерий границы верхушки ветви — ее окраску. Верхушки роста всегда бывают светлее, чем основная часть тела колонии. Более светлая окраска объясняется незаселенностью этой зоны зооксантеллами. Чем быстрее идет продольный рост, тем протяженнее светлая зона, тем длиннее дистальная часть ветви, которую предлагается называть верхушкой. Окраска—достаточно точный показатель локализации и интенсивности зоны роста, а следовательно, и границ верхушки ветви. Обычная длина верхушки 5—10 мм.
Ветви колонии характеризуются, как правило, несколько замедленной кальцификацией. В них продолжается рост скелетных элементов в ширину, из-за чего полости внутри скелета со временем становятся меньше. Одновременно происходит надстраивание элементов скелета на периферии ветви, т. е. постепенное увеличение ее диаметра.
На поверхности ветви могут формироваться радиальные кораллиты 2-го порядка (подразделение на радиальные кораллиты 1-го и 2-го порядков вводится впервые). Радиальные кораллиты 2-го порядка (рис. 1, A, 3) отличаются несколько меньшими размерами, более округлой формой устья, иногда червеобразно изогнутой формой тела и, самое главное, в отличие от радиальных кораллитов 1-го порядка, их ориентация на поверхности ветви может быть практически любой (тогда как радиальные кораллиты 1-го порядка лежат всегда в одной плоскости с ветвью и ориентированы устьями только дистально). Морфологические различия радиальных кораллнтов определяются их происхождением: кораллиты 1-го порядка формируются на верхушке роста на базе аксиального кораллита, а радиальные кораллиты 2-го порядка образуются на остальной части ветви на базе радиальных кораллитов 1-го порядка.
У многих видов акропор рост радиальных кораллитов в проксимальных частях ветвей и особенно на их обращенных к субстрату сторонах настолько затормаживается, что скелет, формирующийся под ценосарком, начинает медленно обрастать кораллиты и в ряде случаев погребать их. Выглядит это как уменьшение длины кораллитов.
В период размножения в ветвях колонии происходит формирование половых продуктов, а затем, после оплодотворения, вынашнвание планул в гастральных полостях полипов. Верхушки же ветвей в размножении не участвуют, что служит дополнительным критерием подразделения ветви на самою ветвь и ее верхушку. В энтодерме зооидов и ценосарка ветви, особенно вблизи поверхности, находится множество зооксантелл.
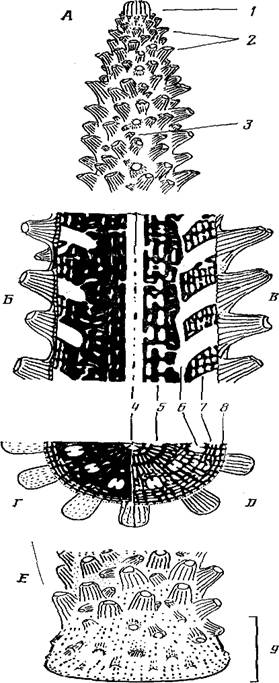
Рис. 1. Схема строения колонии Асrороrа formosa: A — дистальная часть ветви, включающая верхушку ветви; Б — Д — строение скелета и системы полостей на продольном (Б—В) и поперечном (Г— Д ) сечениях в дистальной и проксимальной (В, Д) частях; Е—основание колонии; / — аксиальный кораллит с формирующимися на боковых ребрах радиальными кораллитами 1-го порядка; 2—радиальные кораллиты 1-го порядка; 3—радиальные кораллиты 2-го порядка; 4 — аксиальный канал; 5 — подсистема циркумаксиальных полостей; 6 — полости радиальных кораллитов; 7— подсистема интеррадиальных полостей; 8 — подсистема поверхностных полостей; 9 — зона инкрустации
Ствол колонии. Ствол и ветви колонии морфологически и функционально похожи. Стволу колонии, как и ветвям, присущи: 1) кальцификация внутренних полостей в порядке от центра к периферии ствола; 2) наличие у ряда видов радиальных кораллитов 2-го порядка; 3) уменьшение размеров радиальных кораллитов вследствие их обрастания скелетом, формирующимся под ценосарком; 4) участие в половом размножении; 5) наличие массы зооксантелл в энтодерме. Отличительная особенность ствола — срастание между собой его ветвей, в результате чего образуется более мощная структура. Форма ствола из-за слияния с ветвями отлична от исходно цилиндрической. Скелет в местах слияний отчасти напоминает скелет в зоне инкрустации. Места срастания характеризуются повышенными кальцификацией и скоростью роста. Формируются как бы дополнительные зоны роста. Ствол колонии выполняет функцию опоры. Если ветви обычно легко отломить, то ствол сломать во много раз труднее, а без специальных инструментов и вовсе невозможно. По-видимому, в стволе происходит перекристаллизация скелета, которая приводит к усилению его прочности.
Основание колонии или зона инкрустации. По краю контакта ствола с субстратом имеется краевая зона или зона инкрустирующего роста (рис. 1, Е). Здесь осуществляется стелющийся рост колонии по субстрату. Формируется зона роста после метаморфоза планулы, следовательно, она свойственна колонии с момента се возникновения. В зоне инкрустации у многих видов акропор образуются радиальные кораллиты, ориентированные устьями к растущему краю. По ориентации радиальных кораллитов легко установить границу между стволом и основанием колонии (рис. I, E, 9).
Структура скелета радиальных кораллитов и остального скелета в зоне инкрустации обычно несколько отличается от таковой ствола. Подошву зоны инкрустации выстилает скелет, состоящий из особых элементов — шестигранных пластинок, плотно прилегающих друг к другу. Нигде более в нормальной колонии такие элементы не встречались. Благодаря разрастающемуся основанию колония прочнее прикрепляется к субстрату. Зона инкрустации, повторяя рельеф субстрата и проникая в мелкие полости, способствует усилению, прочности связи между колонией и субстратом. В зоне инкрустации возможно образование новых ветвей колонии на основе разрастания отдельных радиальных кораллитов. Для некоторых видов, например A. humilis (Dana, I846), это основной способ ветвления. Еще одна функция зоны инкрустации—предотвращение обрастания колонии другими кораллами. Конкурентная борьба между колониальными кораллами часто осуществляется при соприкосновении их оснований.
В ряде случаев инкрустирующий рост играет важную роль в процессе реколонизации. Нередко вся базальная часть колонии заносится песком и отмирает. После отступления песка выжившая часть колонии формирует зону инкрустирующего роста и обрастает затем свой же отмерший скелет. Так может повторяться много раз, и в строении скелета в соответствующей части колонии это находит ясное отражение. У разных видов Асrороrа соответствующие четыре части колонии развиты не в одинаковой пропорции. Так, A. formosa, A. pulchra (Brook, 1891), A. aspera имеют слабо выраженный ствол, но ярко выраженные ветви. Напротив, у A. hyacinthus (Dana, 1846) и A. clathrata (Brook, 1891) веточки маленькие, а ствол, представляющий продукт слияния ветвей, составляет основную часть тела колонии. У A. humilis и A. digitifera (Dana, 1846) ствол вообще сливается с зоной инкрустации и не может быть от нее отделен. Подобных примеров много, они требуют специального изучения. Для осуществления подобной работы необходим сбор целых колоний, а не нх фрагментов, при точном указании глубины, особенно вблизи поверхности рифа. Применение аквалангистской техники позволяет это осуществить.
Система полостей в скелете колонии
Многочисленные разноразмерные полости в скелете ветви Асrороrа расположены довольно упорядоченно. В размещении полостей прослеживается радиальная симметрия. Кроме того, полости расположены концентрическими окружностями (рис. 1, Г). Более подробное изучение системы полостей показало, что все они связаны между собой, но могут быть подразделены на пять основных подсистем.
В центре по оси ветви проходит аксиальный канал (рис. 1, А,Б,Г, 4). Полость его поделена шестью продольными септами на секторы. Аксиальный канал представляет продолжение гастралыюй полости аксиального кораллита, мезентерии которого простираются по каналу на расстояние нескольких сантиметров.
Два концентрических круга мелких полостей (рис. 1, А, Б, Г, 5) окружают аксиальный канал. Так как терминология для этих полостей не разработана, то можно называть нх подсистемой циркумаксиальных полостей. Скелет, окружающий аксиальный канал и эти полости, формируется в процессе роста аксиального кораллнта. Циркумаксиальные полости закладываются по периферии аксиального кораллита и являются таким же его продолжением, как и аксиальный канал.
Более крупные полости, выявленные на сколе то ближе, то дальше от оси ветви, — это полости радиальных кораллитов. В каждой такой полости можно обнаружить пару направляющих септ. Полости различаются на поперечном сколе по размерам: чем они ближе к центру, тем мельче. Это, во-первых, объясняется тем, что полости у радиальных кораллитов расширяются к устью и, следовательно, чем дальше от устья рассматривать сечение полости, тем оно мельче. Во-вторых, плоскость поперечного скола по-разному сечет полость изогнутого радиального кораллита: чем ближе к периферии, тем более касательно.
Полости радиальных кораллитов со всех сторон окружены подсистемой интеррадиальных полостей (рис. 1, Б, Г, 7). Эти полости делают скелет пористым, и благодаря им осуществляется связь между кораллитами. Скелет, в котором расположены интеррадиальные полости, состоит из радиалыю расходящихся стенок с отверстиями. Стенки довольно регулярно соединены между собой поперечными балками.
На поверхности скелета ветви между его ребрами и столбовидными элементами находится подсистема поверхностных полостей, ограниченная с внешней стороны ценосарком (рис. 1, Б, Г, 8). На внешних краях элемента скелета происходит их нарастание (рис. 2, А), что приводит к формированию очередного “этажа” и замыканию полостей, лежавших первоначально на поверхности, т. е. приобщение их к подсистеме интеррадиальных полостей (рис.2, Б). Иными словами, каждая полость в интеррадиальной подсистеме была в свое время поверхностной.
Полости в колонии обычно выстланы живой тканью, что легко обнаружить по окрашиванию ее витальными красителями. Полости служат для прохода мезентериальных нитей (аконций), длина которых может приблизительно в 10 раз превышать длину кораллита. На сколах хорошо видно, как аконции движутся по проходам.
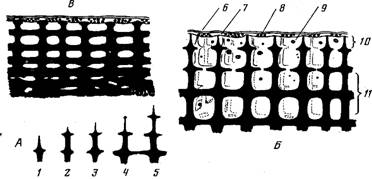
Рис. 2. Схема формирования и строения скелета на участке между кораллитами у поверхности ветви коралла рода Асrороrа, а также взаимное положение скелета и ценосарка (ткани, выстилающие скелет внутри колонии, не изображены): A—схема последовательных этапов роста столбовидных элементов скелета; Б—молодой участок ветви с острыми столбовидными элементами скелета; В—старый участок ветви с тупыми элементами скелета на поверхности и полностью заросшими полостями в центре ветви: /—5 — последовательные этапы роста элемента скелета; 6 — эктодерма ценосарка; 7— энтодерма ценосарка; 8 — зооксантсллы в энтодерме; 9 — мигрирующие клетки и частицы; 10—подсистема поверхностных полостей; П—подсистема интеррадиальных полостей
Очевидно, ткань, выстилающая скелет, не прекращает кальцификацию, так как со временем полости уменьшаются в размерах вплоть до полного их исчезновения (рис, 2, В). Скелет в старых частях колонии слабопористый, почти монолитный (рис. 1, Д). Естественно, что периферические полости в старой части колонии могут быть совсем молодыми и поэтому крупными. Из-за кальцификации центра ветви в старых ее частях транспорт веществ в них может осуществляться только по периферии, и главной в этом отношении становится подсистема поверхностных полостей.
Распределительная система в колонии
Система полостей в колонии определяет морфологическую основу распределительной системы. О распределительной системе у герматипных кораллов в литературе нет сведений. Эксперименты над акропорами позволили внести некоторую ясность по этому вопросу. С помощью инъекций мелких частиц активированного угля (использовались медицинские таблетки угля) удалось установить, что жидкость, заполняющая внутренние полости, находится в движении Сквозь покровы ценосарка и полипов было видно перемещение инъецированных частиц угля. По-видимому, движение внутренней жидкости осуществляется благодаря работе ресничек. Через ротовые отверстия полипов жидкость не выходит наружу колонии, т. е внутренняя среда достаточно замкнута.
Попытки косвенного изучения распределительной системы без повреждения ветви оказались безуспешными, поэтому была разработана простая методика для регистрации внутренних течений при вскрытии колонии. Часть ветви отсекали, переворачивали сколом кверху, и обе половинки укрепляли вертикально в аквариуме. Поверхности скола на обеих частях оказывались обращенными вверх. На каждую поверхность из пипетки наносили раствор витального красителя (нильский голубой). Краситель засасывался в те полости, через которые течение шло внутрь колонии, и окрашивал ткани, выстилающие эти полости. Напротив, там, где направление течения было из ветви, краска этим течением отгонялась, не приходила в соприкосновение с тканью и не окрашивала ее. Результаты эксперимента и его методика показаны на рис. 3.
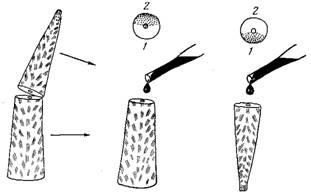
Рис. 3. Схема экспериментальной индикации постоянных течений внутри ветви у акропор с помощью витального красителя: 1,2 — изначально обращенные вверх (/) и к субстрату (2) стороны ветви на схеме поперечного скола. Точками отмечены окрасившиеся участки скола, в которых засасывающее течение было направлено внутрь ветви
Окраска показала, что на двух частях одной ветви окрашивались противоположно расположенные секторы. Это означает, что по одной стороне ветви течение идет к верхушке ветви, а по другой — к ее основанию. В аксиальном же канале преимущественно наблюдалось течение, направленное к основанию. Эксперимент, повторенный с толченым углем, дал такие же результаты. Непосредственное наблюдение под бинокуляром за исходящим течением по движению выносимых им частиц и клеток также подтвердило опыт. Эксперимент был расширен путем инъекции угля в проксимальную часть неповрежденной ветки. Благодаря тому, что в проксимальной части распределительная система ограничена периферическими полостями, здесь можно наблюдать за движением частиц угля под покровами достаточно долго, так как частицы не могут проникнуть в глубь колонии. Результат и этого опыта не противоречил полученным ранее данным. Следовательно, наблюдаемый эффект не является результатом повреждения ветви при ее расколе.
Описываемые опыты были проведены на колониях A. formosa. Затем эффект был проверен и на других видах: A. aspera, A cerealis (Dana, 1846), A. digiiifera, A. millepora (Ehrenberg, 1834), A, nasuia (Dana, 1846), A. pulchra, т. с. на видах с разным типом строения колонии. Везде результат был одинаков. Оказалось, что течение идет к верхушке по стороне ветви, обращенной вверх, а от верхушки к основанию — по стороне, обращенной к субстрату. Переворачивание ветви вокруг собственной оси на 180° не меняло хода течения. Следовательно, течение в распределительной системе определяется в процессе роста колонии и не зависит от положения ветви в эксперименте. Между секторами с дистально и проксимально направленными течениями были пограничные нейтральные зоны, в которых не были зарегистрированы течения.
Интересно, что данный тип распределительной системы — функционально билатеральный — никак не согласуется с радиальной симметрией системы полостей, служащей морфологической основой для распределительной системы. Следует в этой связи отметить, что течения внутриполостной жидкости на своем пути пересекают, не отклоняясь, все полости подряд, и полость кораллита оказывается такой же частью русла потока, как и все остальные полости.
Полученные результаты заставляют несколько изменить традиционное представление о колонии коралла—полипняке—как о продукте бесполого размножения. Колония у акропор, как это следует из приведенных данных, не агрегат зооидов, как-либо оставшихся в связи друг с другом из-за незавершенного почкования. Колония представляет собой безусловное целое с характерной формой, дифференциацией ма части и разделением функций.
Этот вывод может иметь значение и для систематики. Дифференциацию тела колонии на морфо-функциональные части можно использовать для более детального описания видов акропор, систематика которых до сих пор несовершенна. Давно известно (Яковлев, 1964; Wood-Jones, 1910), что герматипные кораллы, особенно акропоры, отличаются пластичной изменчивой формой, что затрудняет их систематику. Однако представление о вариабельности формы колонии во многом связано, во-первых, с отсутствием исследований морфологии колонии в целом, а во-вторых, с недостатком экспериментальных исследований зависимости формы колонии от генотипа и факторов внешней среды. Лишь после осуществления таких исследований можно будет делать выводы о степени пластичности формы колонии и о четкости границ между видами акропор.
Необходимо подчеркнуть, что скелет у акропор, хотя и наружный по своему первоначальному происхождению, но архитектонически безусловно внутренний, и все функции, им выполняемые, соответствуют таковым внутреннего скелета.
Приведенное выше описание морфологически и функционально оформленной распределительной системы у кораллов ставит задачу изучения характера и степени интеграции их колонии. Следует признать наличие некоей физиологической интеграции в колонии у акропор, хотя мы еще далеки от понимания значения и специфики регуляторных процессов в ней.
Список литературы
Яковлев Н, Н, I964. О некоторых результатах исследований коралловых рифоз Индийского океана и Красного моря. В кн.: Организм и среда. М.—Л.: Наука, 42—50. Lamberts A. E; 1974.
Measurement of alizarin deposited by corals.—In: Proc. II Intern.Coral Reel Symp., 2. Brisbane: Great Barrier Reef Comittee, 2, 241—244— 1978.
Согаl growth: alizarin method.— In: Coral reefs: research methods. UNESCO, 523—527. Wallace С. С., 1978.
The coral genus Acropora (Scleractinia: Astocoenina: Acroporidae) in general and Southern Great Barrier Reef province.—Mem. Qd. Mus„ 18, 2, 273—319. Wood-Jones F., 1910. Corals and atolls. London, 1—392.