


Исследование коллекции вида пшеницы Triticum spelta L. по полиморфизму глиадинов
Ю.А. Романова, Н.К.Губарева, А.В.Конарев*, О.П.Митрофанова, О.А.Ляпунова, Н.А.Анфилова, П.П.Стрельченко
С использованием запасных белков эндосперма зерновки - глиадинов как маркеров дана оценка генетического разнообразия 170 образцов коллекции пшеницы спельта, собранной в ВИРе. Выявлен широкий внутривидовой полиморфизм по электрофоретическим спектрам глиадина. По степени сходства спектров выделены группы образцов и осуществлена генетическая структуризация коллекции.
Triticum spelta L. (пшеница спельта) - гексаплоидная пшеница с геномным составом, гомологичным таковому мягкой пшеницы. К.А.Фляксбергер (1) и П.М.Жуковский (2) характеризуют спельту как весьма неприхотливую культуру, произрастающую на бедных почвах в гористых местностях. Она относительно устойчива к холоду и избыточному увлажнению.
Повышенное внимание к спельте во многих странах Европы в последнее десятилетие обусловлено рядом причин, среди которых можно назвать пригодность для низко затратного земледелия, весьма популярного во многих развитых странах, а также некоторые пищевые и технологические достоинства, позволяющие ей в ряде случаев потеснить традиционно доминирующую мягкую пшеницу (3-5). Для спельты характерно повышенное содержание белка в зерне -до 21% (6). Мука, полученная из зерна этой пшеницы, пригодна для изготовления лучших по качеству кондитерских изделий (3-5).
Происхождение спельты окончательно не ясно (1,7,8). Сообщалось о произрастании этой культуры не только в Европе, но и в Иране, Закавказье, Центральной Азии, что не позволяет исключить независимое происхождение этой пшеницы в указанных регионах.
Коллекция спельты в ВИРе насчитывает 170 местных и селекционных сортов (образцов), представляющих все основные регионы возделывания и разнообразия этой пшеницы. Коллекция начала формироваться с 1910 г. В основном она включает материалы экспедиционных сборов отечественных и зарубежных ученых, а именно Н.И.Вавилова в Германии и Испании в 1927 г., Н.Н.Кулешова в Швейцарии в 1928 г., H.Kuckuck в Иране в 1952-1954 гг., В.Ф.Дорофеева в Закавказье в 1961 г., Р.А..Удачина в Центральной Азии в 1965-1971 гг. Часть образцов получена по выписке из учреждений различных стран.
Наше исследование было предпринято с целью изучения генетического разнообразия коллекции спельты с использованием электрофоретического полиморфизма запасных белков семян - глиадинов. Такой подход уже более 30 лет успешно применяется во ВНИИР им. Н.И.Вавилова для решения различных фундаментальных и прикладных проблем генетических ресурсов растений (9). Среди них наиболее актуальными являются проблемы структуризации генетического разнообразия и рациональной организации коллекций, включающей идентификацию и регистрацию образцов, контроль за их подлинностью и сохранностью генетической конституции, а также выявление дублетных и генетически очень близких образцов. Эти направления рассматриваются в настоящее время как приоритетные в деятельности научных центров по сбору, сохранению и изучению генетических ресурсов растений и генных банков мира (10,11). В работе с генетическими ресурсами растений весьма эффективным оказалось использование методов молекулярной биологии (9-12).
С учетом всего сказанного выше были поставлены следующие задачи:
1. изучить внутривидовой полиморфизм пшеницы спельта по запасным белкам - глиа-динам на материале коллекции ВИР;
2. выяснить степень родства по спектрам глиадинов между образцами спельты различного географического происхождения и получить новую информацию о генетической структуре коллекции этой культуры для более эффективного использования генофонда спельты в селекции.
Материалы и методы
Метод электрофоретического анализа глиадинов и принципы использования полиморфизма запасных белков в анализе генетических ресурсов злаков были описаны неоднократно (9,12). Материалом для исследований послужили 170 образцов пшеницы спельта. Образцы характеризовали по глиадиновым спектрам путем позернового анализа. Размер случайной выборки составлял 20-100 зерновок на образец. Спектры глиадина записывали в виде белковых формул в соответствии с эталонным спектром (9). В тех случаях, когда компоненты были представлены двумя или тремя субкомпонентами, для обозначения последних использовали нижний индекс при сохранении основной позиции компонента. Например, наличие, соответственно, двух и трех субкомпонентов в позициях а 6 и w4 записывали в формулах как a6j62 и w4j4243.
На основе полученных электрофоретических спектров глиадина была составлена компьютерная матрица исходных данных, в которой наличие компонента обозначали как 1, а отсутствие - 0. Дальнейшую обработку результатов проводили двумя независимыми методами многомерной статистики: кластерным анализом и методом главных компонент. На основе матрицы исходных данных рассчитывали матрицу коэффициентов сопряженностей по Dice (13). При этом величина коэффициента, равная единице, свидетельствовала об идентичности образцов по спектрам глиадина. Расчет матрицы коэффициентов сопряженностей, кластерный анализ (алгоритм UPGMA) и построение фено-граммы проводили с использованием программного обеспечения NTSYS 2.0 (14).
Метод главных компонент (Q-техника) был применен с использованием программного обеспечения STATISTICA 5.0 (15) для прямой классификации изученных образцов по данным анализа спектров глиадина. Для этого матрицу исходных данных транспонировали и использовали для расчета корреляционной матрицы, которая служила основой при выделении главных компонент варьирования массива данных и вычисления величин факторных нагрузок изученных образцов по каждой компоненте.
Результаты
Общая характеристика спектров глиадина пшеницы спельты.
Суммарный спектр глиадина спельты, как и мягкой пшеницы, состоит из a-, b-, g- и w-зон (или фракций), содержащих, соответственно, в первых двух зонах от 3 до 5 компонентов, в д— от 2 до 4 и в w - от 5 до 7 компонентов. Позерновой электрофоретический анализ глиадина установил наличие у спельты значительного внутривидового полиморфизма по данному признаку, причем различия по спектрам глиадина выявлялись не только между образцами, но и между генотипами внутри образцов.
Разные типы спектров формировались как за счет наличия-отсутствия компонентов и субкомпонентов в различных фракциях, так и за счет варьирования их интенсивности. В а- и Ь-фракциях было выявлено, соответственно, по 8 и 10 различных комбинаций компонентов, в д-фракции - 12 комбинаций, из которых наиболее часто встречаемыми были 2t34, 2j35, 2334. В w-зоне найдено более 30 различных комбинаций компонентов. Для всех вариантов w-зоны присущи компоненты в позициях 8 и 9, маркирующие хромосому Ш (9). Наиболее часто встречались сочетания 8292 и 8j8292, реже S^^lOj. Для спельты в w-зоне характерны также комбинации компонентов 34р 4j43, 242. В целом для 170 образцов спельты выявлено 116 типов спектра. В табл.1 приведены примеры белковых формул глиадина образцов спельты различного происхождения.
Характеристика коллекции спельты по спектрам глиадина.
Анализ всего разнообразия спектров глиадина спельты показал, что 71 образец характеризовались одним специфическим для каждого образца типом спектра глиадина и были идентифицированы как мономорфные. Для каждого из остальных 99 образцов было выявлено от двух до восьми типов спектра глиадина (биотипов). Мономорфные и полиморфные образцы имели широкое географическое распространение. Полиморфные преобладали среди образцов Испании, Ирана, Азербайджана и Таджикистана. Некоторые мономорфные и полиморфные образцы имели идентичные или очень близкие спектры глиадина (табл.2). Среди мономорфных образцов выделено семь групп с идентичными спектрами (табл. 2, №№ 1 -7), среди полиморфных - две группы (табл.2, №№8 и 9). Были обнаружены также мономорфные образцы, которые имели спектр глиадина, идентичный одному из типов спектров глиадина некоторых полиморфных образцов (табл.2, №№10-13). Найдены также полиморфные образцы (табл.2, №13), не различающиеся между собой по доминирующему типу спектра (встречаемость более 90%). Такие мономорфные и полиморфные образцы нами были отнесены к генетически близким образцам. Наиболее часто мономорфные образцы с идентичными спектрами встречались среди образцов Германии, а полиморфные генетически близкие - среди образцов из Таджикистана.
Сравнение спектров глиадина образцов спельты разного географического происхождения по составу компонентов и их интенсивности, встречаемости определенных комбинаций компонентов позволило выявить определенную специфику в составе глиадинов. Для 86 образцов из Германии и других стран Европы идентифицировано 42 типа спектра. К характерным особенностям этих спектров относятся наличие в большинстве случаев в а-зоне компонентов 6j62715 а также высокая встречаемость в g-зоне компонентов 2t34 и 2334.
В отличие от предыдущей группы образцы из Испании и Франции давали 12 типов спектра со следующими специфическими комбинациями компонентов: a- 246j62715 g- 2t35, 2235, 12235, w-4j42 или 4j435. Для восьми образцов из Ирана и Алжира выявлено девять типов спектра глиадина. Эта группа характеризовалась наличием а-компонентов вхв31х12. Для шести образцов из Азербайджана идентифицировано шесть типов спектров. Характерных только для этих образцов сочетаний компонентов выявить не удалось.
Образцы спельты из Центральной Азии (Таджикистан, Узбекистан и Туркменистан) выделялись наличием компонентов a3627j72 и wS^^IOj. Внутри группы выявлено значительное разнообразие по электрофоретическим спектрам глиадина: для 44 образцов - 21 основных типов спектра, из которых только 10 специфичны для отдельных образцов. Остальные образцы могут быть объединены в группы, состоящие преимущественно из двух-трех образцов.
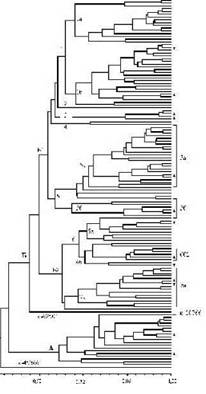
Полученные результаты по разнообразию спектров глиадина были обработаны методами кластерного анализа и главных компонент. На рисунке представлена фенограмма кластеризации образцов спельты. Фенограмма состоит из двух крупных кластеров (рис., А и Б), со степенью родства соответствующей коэффициенту сопряженности по Dice, равному 0,49. Кластер А сформировали 28 образов спельты происхождением из Таджикистана (Dice=0,65). Исключение - образец к-20766 из Германии. Кластер Б объединил все остальные образцы и состоял из двух крупных соподчиненных подкластеров (рис., Б-1 и Б-2), в каждом из которых наряду с образцами из европейских стран присутствовали образцы азиатского происхождения. Образцы к-45366 из Азербайджана и к-62501 из США не вошли ни в один из кластеров.
Внутри каждого кластера и подкластера могут быть выделены более мелкие группы, объединяющие образцы в большей степени близкие друг к другу вплоть до групп образцов с идентичными спектрами глиадина. Образцам с идентичными спектрами глиадина соответствует коэффициент по Dice равный единице. Такие образцы на фенограмме занимают одну позицию, которая помечена звездочкой. В кластере А такие позиции соответствуют 11 и четырем образцам. В кластере Б идентифицировано 11 таких групп, объединяющих от двух до девяти идентичных по спектрам образцов (рис.). Эти образцы могут быть дублетными или генетически очень близкими.
Группа 1 подкластера Б-1 (Dice=0,68) объединила 50 монотипных образцов, а также 12 биотипов полиморфных образцов (рис., группа 1). Подгруппа 1а включила восемь образцов из Германии, по два- из Швейцарии и Югославии и по одному из Польши, Украины и США. Подгруппа 16 также оказалась состоящей преимущественно из образцов европейского происхождения. Однако в этой подгруппе оказались три образца из Таджикистана и по одному из Ирана и Алжира. Два образца из Таджикистана на фенограмме занимали ближние к группам 3 и 4 позиции (рис.). Группы 3 и 4 объединили 11 образцов из Азербайджана, Таджикистана и Туркмении. Цифрой «2» на фенограмме обозначено положение образца к-20379 из Германии. Группа 5, как и группа 1, также объединила образцы преимущественно из европейских стран. При этом 12 образцов из Испании расположились на фенограмме компактно в составе подгруппы 5а (рис.). Вместе с образцами испанского происхождения подгруппа 5 а включала по два образца из Франции, Чехословакии (до 1992 г.) и Швейцарии и один - из Германии (биотип 2 образца к-1734). Подгруппа 56 объединила 9 образцов из европейских стран - Германии, Чехии, Франции, Дании, Швейцарии.
Группа 6 подкластера Б-2 объединила 24 образца (рис.). Подгруппа 6а оказалась сборной. Она содержала образцы из Германии (5 образцов), Азербайджана (2), Австрии (1) и Швейцарии (1). Подгруппа 66 состояла из образцов азиатского происхождения: шесть образцов из Ирана (рис., 662), четыре из Таджикистана и один из Марокко. Подгруппа 7а объединила образцы только европейского происхождения: из Германии, Австрии, Бельгии, Швейцарии и Украины.
Отдельно следует сказать про позиции на фенограмме разных биотипов одного полиморфного образца. Для того чтобы получить реальную картину взаимоотношений в коллекции маркируемых спектрами глиадина биотипов спельты, они были включены в базу данных как самостоятельные образцы. На фенограмме разные биотипы одного образца нередко располагались в разных группах или даже разных подкластерах.
Расчет факторных нагрузок образцов пшеницы спельта методом главных компонент по данным анализа спектров глиадина показал следующее. При обработке исходной матрицы данных было выявлено, что вся исследованная совокупность образцов разделилась на восемнадцать групп, восемь из которых по количеству входящих в них образцов можно отнести к основным группам, обозначенным, соответственно, «Компоненты 1-8» (табл. 3). К одной главной компоненте относили образцы, у которых значения факторных нагрузок были равны или превышали 0,5. Нагрузки меньшей величины не учитывались. Если образец имел одинаковые по величине факторные нагрузки одновременно более чем с одной главной компонентой, его относили к компоненте более низкого порядка. Табл. 3 дает представление о составе выделенных групп.
Группа «Компонента 1» включала 15 образцов (точнее - биотипов), из которых семь - из Германии, а остальные - из Югославии, Швейцарии, США и Польши. По составу входящих образцов она соответствовала подгруппе 1а на фенограмме. Почти равная по численности группа «Компонента 2» состояла из образцов таджикского происхождения и образца к-20766 из Германии. Эта группа соответствовала кластеру А. Группа «Компоненты 3» объединяла 11 образцов из Испании, два - из Франции и по одному из Германии и Чехословакии (до 1992 г.). Некоторые образцы этой группы обнаруживали также одновременно принадлежность к «Компоненте 4». Эта группа соответствовала подгруппе 5 а на фенограмме. Группа «Компоненты 4», включающая 19 образцов, имела неоднородный по происхождению состав (табл.3). Такой же разнородной была и группа «Компоненты 5» (23 образца). На фенограмме этим группам соответствуют подгруппа 16 и группа 6. Четыре образца «Компоненты 6» (табл.3) были идентифицированы в составе группы 5а (рис.). Образцы «Компоненты 7» составили группу 3 (рис.), а образцы «Компоненты 8» - подгруппы 1а, 16 и 6а (рис.).
Обсуждение
Суммируя все сказанное выше, можно сделать вывод о значительном внутривидовом полиморфизме спельты по спектрам проламинов, или по глиадинкодирующим локусам. Как было показано во многих работах (16,17), полиморфизм по спектрам глиадинов мягкой пшеницы обусловлен генетическими различиями по шести мультигенным семействам (Gli-локусам), расположенным в хромосомах первой и шестой гомеологичных групп. Считается, что семейство глиадин-кодирующих генов возникло путем последовательных множественных дупликаций и дивергенции предковых генов внутри мультигенных семейств, их рекомбинации (18). Существование множества аллелей генов мультигенного семейства служит основой того, что каждый генотип может иметь характерную только для него комбинацию глиадинкодирующих генов и, как следствие, уникальный компонентный состав глиадина. Есть все основания считать, что обнаруженный в коллекции спельты полиморфизм по глиадиновым спектрам свидетельствует о различии их по специфическим наборам аллельных комбинаций генов мультигенных семейств. С одной стороны выявлено генетическое разнообразие образцов коллекции спельты, а с другой - их генетическая неоднородность.
Для получения более объективной характеристики структуры генетического разнообразия коллекции спельты мы применили два метода многомерной статистики - кластерный анализ и метод главных компонент, которые хорошо зарекомендовали себя в решении аналогичных проблем биологии. Сравнительный анализ спектров глиадинов этими методами позволил объединить образцы в группы по степени их генетического сходства. Наиболее четко выделились группы образцов, происходящих из Германии и ряда европейских стран, а также из Испании и Таджикистана (табл. 3, рис.). Образцы из Ирана не образовали самостоятельной группы по данным метода главных компонент. В тоже время на фенограмме шесть иранских образцов образовали подгруппу (рис., 662) и оказались расположенными в одной группе с образцами из Таджикистана и Морокко (рис, 66). Образцы из Азербайджана в одном случае попали в группу с образцами из Таджикистана, а в другом - из Германии. Один образец (рис., к-45366) не вошел ни в один из кластеров. По данным метода главных компонент три азербайджанских образца образовали отдельные группы («Компоненты» 9, 12, 17). Остальные образцы вошли в другие группы как и по данным кластерного анализа.
Выделенные нами группы в основном соответствуют систематическому и эколого-географи-ческому делению пшеницы спельта (1,2,7,8). Так В.Ф.Дорофеев и др. (8) в пределах вида T.spelta L. различают по комплексу признаков европейскую (ssp. spelta) и азиатскую (ssp. kuckuckianum) спельты. В пределах первого подвида ими выделены две эколого-географические группы - баварская (proles bavaricum), в основном представленная образцами, собранными в Германии и Швейцарии, и иберийская (proles ibericum), содержащая образцы из Испании. По мнению этих исследователей (8) именно баварская спельта широко распространилась в другие страны Европы (Австрия, Бельгия, Швеция, Испания, Франция, Италия и др.). Распространение иберийской спельты ограничено Испанией (8). Азиатский подвид деления на эколого-географические группы не имел (8). Представителей этого подвида находили в Иране, Азербайджане, Таджикистане и Туркмении, редко в Армении и Грузии. В соответствие с концепцией происхождения спельты, выдвинутой П.М.Жуковским (2), предполагается независимое ее возникновение в регионах: Астурия (Испания), Альпийские горы (Швейцария, Германия), Иран, Азербайджан, Таджикистан и Туркмения.
Полученный нами экспериментальный материал представляет значительный интерес для уточнения внутривидовой классификации спельты. Выделенные по спектрам глиадина германская и испанская генетические группы соответствуют упомянутым эколого-географическим группам европейского подвида. Образцы коллекции ВИР баварской спельты по данным анализа полиморфизма глиадинов представлены одной небольшой и тремя более крупными по числу представителей генетическими группами. Выявленная дифференциация баварской спельты может послужить основой для осуществления более детальной эколого-географической и таксономической классификаций этого подвида. По данным электрофореза глиадинов нами выявлена дифференциация и азиатской спельты. По-видимому, можно ставить вопрос о выделении в составе азиатского подвида также нескольких генетических групп.
В настоящее время одним из приоритетных направлений деятельности генных банков является формирование так называемых оптимальных коллекций, характеризующихся высокой степенью генетического разнообразия при минимальном дублировании материала. Нами были выявлены образцы, которые вполне могут быть отнесены к дублетам или к «генетически очень близким образцам». После дополнительного сравнительного изучения по другим признакам и в случае подтверждения дублетной природы таких образцов, они могут быть переведены в ранг резервных с более редким циклом репродукции.
Особого внимания требует организация работы по сохранению генетической конституции как мономорфных, так и полиморфных образцов. Поскольку в процессе репродукции необходимо воспроизводить все генотипы в тех же количественных соотношениях, как и в оригинальном образце, целесообразно использовать объективные и эффективные маркеры генотипического (био-типного) состава образцов. То, что разные биотипы одного образца иногда обнаруживались в разных группах свидетельствует о том, что уровень генетических различий внутри образцов может превышать уровень различий между образцами. Существование биотипов внутри образцов может быть обусловлено их генетической природой (9,12,16) или быть следствием механического засорения при неоднократном воспроизведении образцов.
Полученные данные по полиморфизму глиадина образцов коллекции спельты могут быть полезны и для уточнения географического происхождения образцов. Особенно это важно для образцов или их биотипов, характеристики которых по спектрам глиадина не совпадают с таковыми большинства образцов данной эколого-географической принадлежности. Использование эффективных маркеров позволяет не только установить сам факт несоответствия, но и получить предварительную информацию о наиболее вероятном происхождении материала. Все рассмотренные аспекты работы с коллекцией чрезвычайно важны для формирования оптимальных коллекций, выяснения генетической структуры коллекции и для унификации системы паспортизации образцов и т.д. Конечным результатом такой работы будет повышение эффективности использования генетических ресурсов (коллекций) в селекции.
Таблица 1. Типы спектров глиадина пшеницы спельты
| Номер по каталогу ВИР | Пр оисхождение | Фракции и компоненты глиадина | Встречаемость C%) | |||
| а. | P | 1 | и | |||
| 1724 | Германия | 5 6i627i | 12 3^4 5i5j | 2i 34 | 2 42 _6a 7i SiSab | 100 |
| 20384 | 324_ 5, | 1 2334 | 34! 62 Ъ Ш | 88 | ||
| 5fii Ъ | 12 324 Jj | 2i 15 | 4i435 71S1S392 | 8 | ||
| 6ф2 7j | 2 324 51 | 2334 | 4i 6; 83?з | 4 | ||
| 24701 | Швейцария | 6i 7i | 123i 4 525j | 12i 34 | 3 425 63 71^3293 | 100 |
| 20539 | Испания | 24 6i62 7i | 12 324 5з | 1 22 3 5 | 4i435 S292 | 87 |
| 24 6i6i 7i | 12 324 5j | 2j 3 5 | 4i4j S 9j | 13 | ||
| 45813 | Иран | 61 637!72 | 2 3j1 5, | 2i 34 | 3 S^iSjS^j | 55 |
| 61 637j72 | 2 324 52 | 2i 34 | 3 42 63 81 92 | 45 | ||
| 45366 | Азербайджан | 5 62 7i72 | 12 Mi5i5253 | 2;34 | 23 63 829j | 100 |
| 52440 | Таджикистан | 45 62 7i | 1 3i3^4 5i52 | 2j22 34 | 2 42 636^2 8292 | 97 |
| 52443 | 4 61162 7i72 | 12 324 52 | 12i 34 | 2 4! 6&Ъ Si 9i93Ю2 | 100 | |
Таблица 2. Образцы спельты с идентичными или близкими спектрами глиадина
| № группы | Номер по каталогу ВНР (происхождение) |
| Идентичные мономорфные образцы | |
| 1 | к-17 24 (Германия) =к-1727 (Германия) =к-1941 (Германия) = к-19097 (Шв enuapi 1я) |
| 2 | K-l^iS (Германия) = к-9991 (Германия) = к-203 -6 (Германия) |
| 3 | K-2O3S2 (Германия) =k-2OJ>S3 (Германия) =к-2ОЗР2 (Германия) |
| 4 | к-20374 (Германия) =ы-2О394 (Германия) =к-40830 (Германия) |
| 5 | к-2469" (Швепцар11я)=к-24 01 (ТЕ1ве11цар11я)=к-2<э343 (Швейцария) |
| 6 | K-2O5GS (Пспанпя)=к-20579 (Испания) |
| 7 | к-24~24 (F.\>гославия) =к-653 5 (Германия) |
| Идентичные полиморфные образцы | |
| S | :|:к-2960~ ((Чехословакия до 1992) =:|:к-1 -34 (Ге|)манггя) |
| 9 | *к-20543 (Испания) ='к-20??8 (Испания) |
| Генетически близкие образцы | |
| 10 | к- 20 "60 (Германия) =' K-4OS2S (Двепига) |
| 11 | к-23383 (Авсцли) ='к-203 80 (Герман1и) |
| 12 | к-56591 (Туркменистан) =:|ъ>52440 (Таджикистан) |
| 13 | к-52443 ^к-524?"7 =|=к-52437 ='=к-52442 =:|1к-52444 =:|ж-52445 =:|1к-5246б (в с е о oj) а дды из Т а дал пд ict ана) |
журнал Генетика, 2001,т.37, № 9, с. 1258-1265.
Таблица 3.Группы образцов T.spelta, выделенные методом главных компонент (Компоненты 1-8) по данным анализа спектров глиадина (образцы в столбцах приведены в порядке уменьшения факторных нагрузок от 0,94 до 0,50)
| Кситпонвст* 1 | Кампюнюста 1 | Кампюнвсга 3 | Кситпонвст* 4 | Иампюнисга 5 | |||||
| NE-бИЕТ. | СП | Ns-биот. | СП | NE-бИЮТ. | СП | NE-бИЕТ. | СП | NE-бИОТ. | СП |
| 6536-1* | ГРМ | 52451-1 | ТАД | 20566-2 | ИСП | 6537-0 | ГРМ | 19372-0 | ЖР |
| 1723-0* | ГРМ | 52460-1 | ££ | 20625-2 | LL | 20764-0 | ££ | 20337-0 | ГРМ |
| 19373-0 | ЖР | 52449-0 | ilil | 20563-0 | ££ | 20032-0 | ££ | 20373-0 | ££ |
| 24608-0 | ШЕЦ | 52469-1 | ££ | 20625-1 | ££ | 20391-2 | ££ | 40329-0 | АЕС |
| 5219-0 | США | 52461-0 | ££ | 20626-2 | ££ | 24696-2 | ШЕЦ | 23552-0 | ££ |
| 15016-0 | ПОЛ | 52468-0 | LL | 20539-0 | ££ | 15015-0 | ПОЛ | 62500-0 | БЕЛ |
| 26340-1 | ШЕЦ | 52447-0 | LL | 21574-0 | ■#РА | 24700-0 | ШЕЦ | 60442-0 | ШЕЦ |
| 200S4-1 | ГРМ | 52438-1 | LL | 20591-0 | ИСП | 20495-0 | ИСП | 23333-0 | АЕС |
| 5218-0 | США | 52448-1 | ££ | 20569-2 | ££ | 24697-0 | ШЕЦ | 45769-0 | ГРМ |
| 24724-0 | ЮГО | 52469-2 | LL | 20541-0 | ££ | 6536-2 | ГРМ | 62503-0 | БЕЛ |
| 46020-0 | ££ | 52451-2 | LL | 20533-1 | ££ | 52471-0 | ТАД | 24703-0 | ШЕЦ |
| 20033-0 | ГРМ | 20766-0 | ГРМ | 1734-2* | ГРМ | 20769-0 | ГРМ | 20330-2 | ГРМ |
| 13294-0 | LL | 52470-0 | ТАД | 45767-0 | ЧЕК | 46136-0 | АЛЖ | 45313-0 | ИРА |
| 20331-0 | ££ | 56569-0 | LL | 20533-2 | ИСП | 45750-0 | ИРА | 24709-0 | ШЕЦ |
| 40331-0 | ££ | 20626-1 | ££ | 45763-0 | ЧЕК | 20374-0 | ГРМ | ||
| 20553-0 | ££ | 24707-0 | ШЕЦ | 40328-0 | АЕС | ||||
| 20546-0 | ££ | 20391-1 | ГРМ | 45317-0 | ИРА | ||||
| 20566-1 | ££ | 24706-0 | ШЕЦ | 24705-0 | ШЕЦ | ||||
| 21573-0 | ■#РА | 20543-0 | ИСП | 52463-0 | ТАД | ||||
| 45814-0 | ИРА | 20377-0 | ГРМ | ||||||
| 21572-0 | ■ФРА | 62502-0 | БЕЛ | ||||||
| 19374-0 | ТЕР | 54441-0 | ТАД | ||||||
| 13296-0 | ГРМ | ||||||||
| Ишппюнисга б | Кампюнюста 7 | Кампюнвсга JS | |||||||
| №-биот. | СП | №-6ивт. | СП | №-биот. | СП | ||||
| 23291-0 | ДАН | 52434-0 | 45369-0 | АЗЕ | |||||
| 29607-0 | ЧЕК | 52452-0 | LL | 35665-2 | ГРМ | ||||
| 19092-2 | ЛАТ | 56591-0 | LL | 1723-0 | ££ | ||||
| 24696-1 | ШЕЦ | 52435-0 | ТАД | 39760-0 | ££ | ||||
| 52464-0 | ТАД | ||||||||
| 62504-0 | ГРМ | ||||||||
| 45363-0 | АЗЕ | ||||||||
Условные обозначения, принятые в таблице:
№-биот. - номер образца по каталогу ВИР - биотип; * - 0, 1,2- биотипы, маркируемые спектрами глиадина;
СП - страна происхождения образца; ГРМ - Германия, ШВЦ - Швейцария, ИСП - Испания, ФРА - Франция, ДАН - Дания, БЕЛ - Бельгия, ЧЕХ - Чехословакия до 1992 г., ЛАТ - Латвия, ТАД - Таджикистан, АЗЕ - Азербайджан, ТУР - Туркменистан, АЛЖ - Алжир, ИРА - Иран, УКР -Украина, ПОЛ - Польша, ЮГО - Югославия.
Рис. Распределение образцов спельты на фенограмме по данным анализа полиморфизма глиадинов.
Обозначения - см. пояснения в тексте
1. Фляксбергер К.А. Пшеница - род Triticum L. «Культурная флора СССР». Хлебные злаки. Пшеница. М.-Л.: Госиздательство совхозной и колхозной литературы. 1935. С. 17-404.
2. Жуковский ИМ. Культурные растения и их сородичи. Л.: Колос, 1971.752 с.
3. Jorgensen J.R. and Olsen С. С. Yield and quality assessment of spelt (Triticum spelta L.) compared with winter wheat (Triticum aestivum L.) in Denmark // In: «Spelt and Quina» - Working Group Meeting, 24-25 October 1997, Wageningen, the Netherlands. 1997. P. 33-38.
4. Eltun R. and Aasven M. The possibilities for spelt cultivation in Norway // In: «Spelt and Quina» - Working Group Meeting, 24-25 October 1997, Wageningen, the Netherlands, 1997. P. 7-13.
5. Dahlstedt L. Spelt Wheat (Triticum aestivum ssp. Spelta (L.)): An alternative crop for ecological farming systems // In: «Spelt and Quina» - Working Group Meeting, 24-25 October 1997, Wageningen, the Netherlands, 1997. P. 3-6.
6. Каталог образцов мировой коллекции ВИР с характеристикой содержания белка и аминокислот. Л.: ВИР. 1972. Вып. 100. 110 с.
7. E.S.McFadden, E.R.Sears. The origin of Triticum spelta and its free-thresching hexaploid relatives //Jounal of Heredity. 1946. V. 37. №3. P. 81-89, № 4.107-116.
8. Культурная флора. Пшеница. Под ред. В.Ф.Дорофеева и О.Н.Коровиной. Л.:Колос. 1979. 346 с.
9. Молекулярно-биологические аспекты прикладной ботаники, генетики и селекции (ред. акад.В.Г.Конарев). Теоретические основы селекции. Т. I. M.: Колос, 1993. 447с.
10. Hodgkin Т. Some current issues in the conservation and use of plant genetic resources // Molecular genetic techniques for plant genetic resources. Report of an IPGRI Workshop. October 1995. Rome, Italy. 1997. P. 3-10.
11. Конарев А.В. Использование молекулярных маркеров в работе с генетическими ресурсами растений // Сельскохозяйственная биология. 1998. № 5. С. 3-25.
12. Yl.Gubareva N.K. and Gaydenkova N.V. Varietal identification and registration of bread wheat genefond by means of gliadin electrophoresis // In: III Int. Sympos. ISTA «Biochemical identification of varieties». L., USSR. 1988. P. 131-134.
13. DiceL.R. Measures of the amount of ecologic association between species//Ecology. 1945. V. 26. P. 297-302.
14. Rolf KG. NTSYS-pc. Numerical taxonomy and multivariate analysis system, version 2.02. Exeter Software, New York. 1995.
15. Statistica for Windows (Computer program manual) // Tulsa, OK: StatSoft, Inc., 2325 East 13th Street, Tulsa, OK 74104, USA. 1995.
16. Созинов А.А. Полиморфизм белков и его значение в генетике и селекции. М.: Наука, 1985. 272 с.
17