


Генотипическое и фенотипическое разнообразие населения Украины по группам крови
Группы крови формируются в результате действия различных эволюционных факторов. Актуальность темы данной работы состоит в том, что группа крови может определять устойчивость к определённым заболеваниям, распределение групп крови территориально может указывать на такие генетические процессы как дрейф генов, миграции.
Целью данной работы является исследование генетической структуры (полиморфизма) украинских популяций по локусам групп крови.
Задачи:
- установить фенотипическую, генотипическую структуру популяций разных областей Украины по группам крови по системе АВ0 и по системе Rh;
- определить частоты аллелей групп крови в популяциях разных областей Украины по группам крови по системе АВ0 и по системе Rh;
- сравнить и проанализировать полиморфизм на территории Украины.
1. ОБЗОР ЛИТЕРАТУРЫ
1.1 История происхождения территориально-административного деления Украины
Долгие века нынешняя территория Украины была разделены и находилась под властью различных государств. Рассмотрим историю некоторых нынешних городов Украины.
В начале нашего века территория Житомирской области заселялась древними восточнославянскими племенами, север – древлянами, центральная часть и юг – полянами. В 945 году древлянские земли окончательно вошли в состав владений киевских князей – Киевской Руси. В X- начале XII веках район проживания древлян и полян входил в состав мощного древнерусского восточнославянского государства – Киевской Руси, затем в состав Владимиро-Волынского, а с 1199года – Галицко-Волынского княжества. Волынь и Полесье принадлежали к славянской прародине, были колыбелью украинского народа. В первой половине XIII века вместе с другими славянскими землями Житомирщина попала под гнет татаро-монгольских завоевателей. В 1362 году этой территорией завладели литовские феодалы, а после объединения Литвы и Польши в 1569 году в единое государство Речь Посполитую, волынские земли попали под власть польских магнатов. После воссоединения правобережной Украины с Российской империей в 1793 году, территория Житомирщины вошла в состав Волынской и частично Киевской губерний.
Край, где находится современная Черновицкая область, заселялся в эпоху палеолита, о чем свидетельствуют обнаруженные здесь стоянки первобытных людей, возраст которых от 40 до 15 тыс. лет. Северная часть современной области в X-XII веках входила в состав Киевской Руси, позже была частью Галицко-Волынского княжества, которое со временем попало под власть Золотой Орды. В 1345 году венгерские войска выгнали татар с правобережья Днестра и Северная Буковина отошла Венгерскому королевству. В 1359 году Молдавское воеводство, входившее в состав Венгерского королевства, объявило о своей независимости. Образовавшееся Молдавское княжество распространило свою власть на всю территории Буковины. В 1514 году Буковина перешла под власть Турции. После русско-турецкой войны (1768-74гг.) Буковина перешла к Австрии, в составе которой и находилась до 1918 года. Этот период стал временем интенсивного развития региона, в 1849 году было образовано герцогство Буковина в составе Австрии. После распада Австро-Венгерской империи, была попытка присоединения ее к Украине, но армия Румынии оккупировала территорию Буковины. В состав современной области также входит часть исторической области Бессарабии (Хотинский район). Бессарабия – древнерусская земля, входящая в состав Галицкого, а позже Галицко-Волынского княжества, в XIV – XV вв. находилась в составе Молдавского княжества, а в XVI веке южные районы Бессарабии стали турецкими. В 1812 году по Бухарестскому мирному договору, заключенному между Российской империей и Турцией, территория Бессарабии отошла к России, на ее территории была образована Бессарабская губерния, одним из уездов который был Хотинский. В 1918 году Бессарабия была оккупирована Румынией. Территории северо-восточной части Королевства Румынии - Бессарабия и Северная Буковина были аннексированы Советским Союзом и 7 августа 1940 года, после присоединения Северной Буковины и Хотинского уезда Бессарабии к Украинской ССР, была образована Черновицкая область.
Закарпатье, отделенное от остальной Украины естественной преградой – Карпатскими горами с несколькими перевалами - столетиями формировало свою особую историю и культуру, которые выделяют самую западную область страны от других украинских регионов. Первые упоминания о жизни людей на территории области относятся к периоду раннего палеолита. Прошлое Закарпатья неразрывно связано с восточными славянами. Отдельные славянские группы здесь появились в первое тысячелетие нашей эры и были выходцами из Прикарпатья и Волыни. В 896 году начался переход венгерских кочевых племен через Карпаты в Среднее Придунайье, освоение которого длилось на протяжении всего Х века. В Закарпатье, которое далеко находилось от основного центра славянства, еще некоторое время сохранялась власть славянской знати. Однако, начиная с XI века и завершая ХIII веком, Закарпатье постепенно вошло в новообразованное европейское государство – королевство Великая Венгрия и здесь окончательно утвердились венгерские короли. До средины ХХ века Закарпатье входило в состав разных государств, но всегда сохраняло свою славянскую сущность. Некоторое время часть Закарпатья входила в состав Галицко-Волынского княжества. Долгое время вся территория Закарпатья принадлежала венгерским правителям, но в 1541 году центральной Венгрией овладели турки, после чего она была разделена, Закарпатье оказалось поделенным на две части. Впоследствии центральные и восточные районы края вошли в Турецкую империю, а западная часть попала под власть Габсбургов. После 160-летнего турецкого ига, в конце XVII века вся территории Венгрии, в том числе и Закарпатье, перешла под власть Габсбургов. После поражения Австрии в австро-прусской войне 1866 года, было создано двуединое государство - Австро-Венгрия, после распада которого, в 1918 году образовался ряд государств, в том числе и Чехословакия. В 1918-19-х годах западная часть территории края была оккупирована чехословацкой армией, а румынские войска заняли юго-восточную часть. В то же время в Венгрии провозглашается Советская республика, включившая в себя большую часть Закарпатья.
Возле Запорожья исследовано несколько поселений периода позднего палеолита. В VII веке до н.э. в Северном Причерноморье проживали скифы. На протяжении многих столетий территория края была своеобразным коридором, которым проходили разные народы, где скрещивались разные культуры. После татаро-монгольского нашествия в 1237-1240 годах, территория Запорожья на два столетия вошла в состав Золотой Орды. После порабощения Украины литовцами, а затем поляками, на запорожских землях в XV веке появились казаки («вольный человек» в переводе с тюркского). Южно-украинские земли стали одним из центров формирования запорожского казачества, которые осваивали земли у порогов Днепра.
Бердянск – город областного значения в Запорожской области Украины, административный центр Бердянского района. На территории современного Бердянска жили люди еще в глубокой древности, о чем свидетельствуют обнаруженные в урочищах Сладкий Лиман и Ближний Шпиль остатки двух поселений эпохи неолита и ранней бронзы (III тыс. до н.э.). После ликвидации Запорожской Сечи и раздачи казацких земель помещикам многие казаки (около 5-7 тыс.), бежали во владения турецкого султана, который был заинтересован в том, чтобы запорожцы перешли к нему на службу, и разрешил им селиться на пограничной территории под Очаковым. Когда Екатерина II потребовала выдачи беглецов, турки на это не пошли и переселили казаков подальше от границы, на правый берег Дуная и в 1778 году султан официально признал запорожских казаков своими подданными. На турецких землях казаки основали Задунайскую Сечь, которая несколько раз меняла свое местонахождение. По словам украинского историка Михаила Грушевского, жилось казакам в Турции неплохо, только «мучила совесть запорожцев, что приходится помогать бусурманам воевать против христиан», поэтому с Задунайской Сечи убегали отдельные отряды запорожцев к своим единоверцам в Россию. В ответ на массовый уход казаков с Украины в Турцию, русское правительство арестовало бывших войсковых старшин и кошевого атамана Петра Кальнишевского, который более 25 лет провел в заключении в Соловецком монастыре. Однако, когда в 1787 году началась очередная война России с Турцией, российское правительство обратилось за помощью к бывшим казакам, которые оставались на территории Украины, из которых было создано «войско верных казаков», переименованное в Черноморское казачье войско в 1788 году. В русско-турецкой войне 1787-1791 годов черноморским казакам пришлось сражаться с казаками из Задунайской Сечи, т.е. своими бывшими соотечественниками.
Совсем другая ситуация сложилась в начале русско-турецкой войны 1828-1829 года, когда часть «турецких» казаков под Измаилом перешла на сторону русской армии. Из-за этого турки уничтожили Задунайскую Сечь. Из тех казаков, кто решил остаться на территории России, было сформовано Азовское казачье войско, которое поселили между Мариуполем и Бердянском.
История Сумской области неразрывно связана с развитием Украинского государства - территория современной области заселялась в конце позднего палеолита. В VIII веке до н.э.- вошла в состав большого государственного формирования племен скифов – Скифии. В начале нашего века на территории Сумщины появились племена древних славян. В Х веке территория Сумщины входила в состав Киевской Руси, в Липовицкое княжество. В XI – XIII веках Сумщина входила в состав Черниговского и Северского княжеств. В 1360-е годы юго-западная часть нынешней области вошла в состав Великого Княжества Литовского, а затем - стала частью Речи Посполитой. После того, как императрица Екатерина II в 1764 году отменила гетманскую власть на Левобережье, а в 1765 году отменила полковой уклад и самоуправление в Слободской Украине, на Сумщину было распространено административно-территориальное деление Российской империи.
Территория нынешней Черкасской области была заселена в древнейшем периоде каменного века – палеолите. Территория современной области входила в состав тех земель Приднепровья, где формировалось ядро восточнославянских племен, из которых возникла могучая феодальная держава – Киевская Русь. В 1239-40-х годах эти земли были опустошены татарами. Воспользовавшись татаро-монгольским нашествие и феодальной раздробленностью Руси, в XIV – XVI веках территорию современной области захватили литовские и польские феодалы.
2. ПОЛИМОРФИЗМ
Согласно определению, полиморфизм — это существование в популяции двух или более резко различающихся (прерывистых) форм, при котором частота более редкой формы определяется не одним лишь мутированием. Иными словами, полиморфизм — это такая изменчивость в локальной воспроизводящейся популяции, при которой проявляется чётко выраженное или резкое менделевское расщепление.
Такое определение полиморфизма не позволяет относить к нему некоторые типы изменчивости. Оно исключает чисто фенотипическую изменчивость (поскольку это негенетическая изменчивость); оно исключает географическую изменчивость (которой не существует в одной популяции); оно исключает полигенную изменчивость (при которой не происходит расщепления на резко различающиеся классы); и, наконец, оно исключает генетическую изменчивость, обусловленную новыми или повторными мутациями.
На основе разных критериев можно выделять различные типы полиморфизма. Важно различать генетический полиморфизм и хромосомную изменчивость. Генетический полиморфизм — это прерывистая изменчивость по гомологичным аллелям одного и того же генного локуса. Хромосомным называют полиморфизм по типам хромосом, например по половым хромосомам, или по таким перестройкам, как инверсии.
Различают также переходный и сбалансированный полиморфизм. В случае переходного полиморфизма разнообразие носит временный характер: оно наблюдается до тех пор, пока происходит процесс замещения одной формы другой при контролирующем действии естественного отбора. При сбалансированном полиморфизме разные типы представляют собой более или менее постоянные компоненты данной популяции благодаря естественному отбору, который благоприятствует сохранению разнообразия.
Все формы полиморфизма — генетический, хромосомный, переходный и сбалансированный — весьма обычны и широко распространены в живой природе. В популяциях организмов с половым размножением полиморфизм в сущности наблюдается всегда.
Явление полиморфизма подводит нас к концепции генофонда, которая в свою очередь даёт возможность по-иному взглянуть на локальную воспроизводящуюся популяцию. Рассмотрим популяцию, полиморфную по гену A и содержащую аллели A1, A2 и A3. В такой популяции будут возникать диплоидные генотипы A1A1, A1A2, A2A2 и т. п., и обусловленные этими генотипами формы будут представлены в любой выборке особей, однако ясно, что одна из главных черт данной популяции — это генетический полиморфизм, лежащий в основе наблюдаемого разнообразия. Можно сказать, что генофонд такой популяции содержит аллели A1, A2 и A3. Кроме того, эти аллели встречаются в генофонде с определённой частотой; допустим, что их частоты равны соответственно 60, 30 и 10%. Следовательно, популяцию можно описать количественно, используя типы генов, содержащихся в её генофонде, и их частоты.
Следует отметить, что концепция генофонда шире, чем концепция полиморфизма. Генофонд популяции слагается из всех имеющихся в ней генов. Так, генофонд нашей гипотетической популяции может быть полиморфным по гену А, содержать редкий мутантный аллель гена В и быть мономорфным по генам С и D.
Особи, из которых состоит данная популяция, в каждом данном поколении представляют собой различные генотипические продукты гамет, взятых из генофонда этой популяции в предшествующем поколении.
2.1 Полиморфизм по группам крови у человека по системе АВ0
Различия в групповой принадлежности крови зависят от различий в строении определенных сложных углеводных компонентов мембраны эритроцитов; это относится, по крайней мере к группам ABO и Lewis, которые наиболее изучены с химической точки зрения.
Групповые вещества крови представляют собой антигены, т. е. при введении индивидууму, лишенному именно этого антигена, вызывают образование специфических антител. Многие из этих антигенов представляют собой белки, однако большинство близки к углеводным антигенам, описанным у бактерий. Антитела — это белки (иммуноглобулины), которые выделяются в плазму крови особыми антителообразующими клетками. Группоспецифические антитела, используемые для выявления антигенов групп крови, обычно обнаруживаются в сыворотке беременных женщин (иммунизированных антигенами собственного плода) или в сыворотке индивидуумов, которым многократно переливали кровь. Иногда такие антитела возникают «спонтанно» у индивидуумов с особыми группами крови; некоторые же (например, анти-М и анти-N) получают путем иммунизации кроликов эритроцитами человека.
Главный метод определения различий по группам крови — агглютинация. Если отмытые эритроциты смешать с сывороткой, содержащей специфические антитела, то они покрываются белком антител и агглютинируют. Однако некоторые специфические антитела, связываясь с эритроцитами, не вызывают их агглютинации; такие антитела называют неполными.
Наследование различных вариантов групп крови удивительно точно следует законам Менделя. Этим можно воспользоваться при решении вопроса об отцовстве, для различения монозиготных близнецов от дизиготных, в работах по изучению сцепления генов. Многие различия по группам крови встречаются в разных популяциях с высокой и неодинаковой частотой; поэтому они представляют ценность и для антропологии. Более того, многие из них могут быть определены сразу после рождения и впоследствии изменяются лишь при нескольких редких заболеваниях. Вначале казалось, что правило «один ген — один антиген» приложимо и в этих случаях; однако с развитием молекулярной генетики и после описания ряда необъяснимых явлений стала очевидной ограниченность этого правила. Известно, что структурные гены представляют собой участки ДНК, в которых закодированы аминокислотные непоследовательности полипептидных цепей белковых молекул. Поэтому углеводные антигены, выявляемые при тестировании групп крови, не могут быть прямыми продуктами генов. Соответствующие гены, как полагают, контролируют ферменты (гликозилтрансферазы), участвующие в синтезе углеводных цепей. Далее, гены состоят из множества азотистых оснований, способных к мутациям. Возможно, что мутация, затрагивающая тот или иной фермент, приводит к изменению не одной антигенной специфичности групповых веществ крови, когда определенная группа антигенных специфичностей наследуется как целое. Ситуацию, когда определенная группа антигенных специфичностей наследуется как целое, можно теоретически объяснить, постулируя существование ряда тесно сцепленных генов; однако если сцепление столь тесно, что кроссинговер практически никогда не наблюдается, этот постулат остается лишь удобным предположением, не проясняя истинной структуры этих генов. Серологические методы чувствительны и высокоэффективны, но для понимания природы генов они дают нам меньше сведений, чем химия. К сожалению, почти ничего не известно о физиологическом значении групповых веществ крови, а ведь знание их функций позволило бы успешно вести поиски агентов, избирательно действующих на эти вещества.
В 1968 г. Рзйс и Сенджер в своей ставшей классической монографии перечислили 14 различных систем групп крови, т. е. группоспецифических веществ, которые контролируются независимыми генными локусами. Кроме того, существует немалое число антигенов групп крови, которые не варьируют (общие антигены), а также антигенов, найденных лишь в одной или нескольких семьях (личные антигены).
2.2 Система А1А2В0
Эта система была обнаружена первой; в 1900 г. Ландштейнер показал, что сыворотка некоторых индивидуумов вызывает агглютинацию отмытых эритроцитов ряда других индивидуумов. Это открытие позволило разработать метод безопасного переливания крови, а также предоставило в распоряжение генетиков и антропологов набор просто наследуемых вариантов, довольно часто встречающихся в большинстве популяций, легко обнаруживаемых и практически не зависящих от пола, возраста и окружающей среды. Затем лишь в 1924 г. Бернштейн установил тип наследования групп крови системы ABO, существо которого представлено в табл. 1. Эритроциты могут быть носителями антигенов А, или В, или А и В, или же ни одного из них; таким образом, мы получаем четыре фенотипа: группы А, В, АВ и О; наследование определяется тремя аллелями— А, В и О, которые при расщеплении дают 6 генотипов — АА, АО, ВВ, ВО, АВ, 00.
Таблица 1
Генетическая и серологическая характеристики групп крови системы ABO
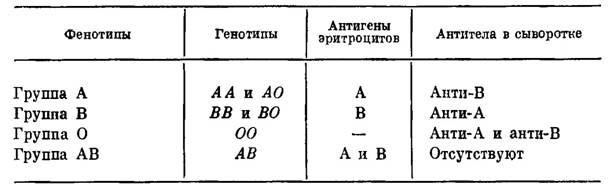
Присутствие антигена А можно определить, смешивая клетки с сывороткой, содержащей анти-А, после чего они агглютинируют; антиген В определяется сходным образом с помощью анти-В. Истинный анти-0 не найден; поэтому мы не можем серологически отличить гомозиготы А или В от гетерозигот А или В (АО и ВО). Отсюда также следует, что мы не можем определить частоты этих генов в популяции путем простого подсчета; для этого приходится применять косвенные методы, исходя из допущения, что в данной выборке для этих аллелей соблюдается равновесие Харди — Вейнберга.
Анти-А, как правило, присутствует в сыворотке индивидуумов, лишенных этого антигена, а анти-В — у тех, кто лишен антигена В. По-видимому, эти естественно встречающиеся антитела образуются в ответ на A-подобные и В-подобные антигены бактерий и других организмов, проникающие в пищеварительный тракт человека вскоре после рождения.
Группу А можно подразделить на группы А1 и А2. Эритроциты А2 слабо реагируют с обычным анти-А, однако у нас нет специфических анти-А2, чтобы идентифицировать этот вариант. Правда, в организме некоторых А2- и А2В-индивидуумов образуются специфические анти-А1,которые можно применить для субтипирования A-клеток. Тем не менее генотипы А1А2серологически неотличимы от А1А1 или А10 , a A2А2 — от A20.Описаны различные другие количественные варианты группы А (например, A-intermediate), довольно часто встречающиеся у африканцев.
Правила переливания крови. Во многих случаях возникает необходимой переливания крови одного человека (донора) – другому (реципиенту). При переливании крови необходимо учитывать агглютиногены донора и агглютинины реципиента. При переливании небольших количеств крови агглютинины донора значительно разводятся и теряют способность агглютинировать эритроциты реципиента. У лиц с I (0) группой крови эритроциты не содержат агглютиногенов (ни α, ни β), такую кровь можно переливать лицам с любой группой крови. Кровь II (А) группы можно переливать только лицам со II (А) или IV (АВ) группой крови (у которых нет агглютининов a). Кровь III (В) группы можно переливать только лицам с III (В) или IV (АВ) группой крови (у которых нет агглютининов b); кровь IV (АВ) группы можно переливать только лицам с IV (АВ) группой, у которых вообще нет агглютининов. Таким образом, лица с I (0) группой крови являются универсальными донорами, а лица с IV (АВ) группой крови – универсальными реципиентами.
Однако изложенные правила пригодны лишь при переливании небольших количеств крови. При необходимости переливания больших количеств крови используется только одногруппная кровь.
2.3 Система Rhesus(Rh)
Резус-фактор — это антиген (белок), который находится на поверхности красных кровяных телец (эритроцитов). Он обнаружен в 1919 г в крови обезьян, а позже — и у людей. Около 85 % европейцев (99 % индейцев и азиатов) имеют резус-фактор и соответственно являются резус-положительными. Остальные же 15 % (7 % у африканцев), у которых его нет, — резус-отрицательный.
Резус-система определяется тремя сцепленными генами (CDE); все эти гены локализованы в 1-й хромосоме. Наиболее сильным антигеном резус-системы является антиген RhD, который контролируется соответствующим геном D. При этом резус-положительная группа крови доминирует над резус-отрицательной.
Наследование резус-фактора происходит сложным образом, но, учитывая ведущую роль гена D, его можно представить как моногенное наследование с полным доминированием: при генотипе DD или Dd резус положительный (Rh+), а при генотипе dd – отрицательный (Rh–). Переливание резус–положительной крови человеку с резус-отрицательной группой крови приводит к развитию иммунной реакции на резус-антиген: синтезируются резус-антитела, то есть происходит иммунизация организма против резус-антигена. Этот процесс продолжается в течение 2…4 месяцев. Резус-антитела сохраняются в сыворотке длительное время. Если иммунизированному человеку перелить резус-положительную кровь, то резус-антитела разрушают резус-положительные эритроциты, что приводит к гемолитическому шоку, а нередко – и к смерти человека.
Резус-конфликт. Большое значение резус-совместимость имеет при вынашивании плода. Если резус-отрицательная женщина (dd) вынашивает резус-положительный плод с генотипом Dd (аллель D приходит от резус-положительного отца DD или Dd), то происходит иммунизация организма матери резус-антигеном.
Существуют различные точки зрения на механизм иммунизации. Согласно одной из них, часть эритроцитов плода попадает в кровоток матери при родах. Однако имеются данные о возможности проникновения резус-антигенов через плаценту еще до родов; в то же время, в некоторых случаях иммунизации не происходит.
Если организм женщины оказался иммунизированным резус-антигеном, то при повторной беременности резус-антитела через плаценту попадают в кровоток плода. Если генотип второго ребенка dd, то резус-антигены отсутствуют, и резус-конфликта не происходит. Но если генотип второго ребенка Dd, то резус-антитела матери разрушают резус-положительные эритроциты плода. Это приводит или к гибели плода, или к рождению неполноценного ребенка.
Таким образом, браки ♀ Rh– × ♂ Rh+ (например, ♀ dd × ♂ Dd или ♀ dd × ♂ DD) оказываются неблагоприятными, т.к. в этом случае велика вероятность рождения неполноценного ребенка. Однако в каждом конкретном случае все может быть не так страшно.
Во-первых, в браках ♀ dd × ♂ Dd (то есть при гетерозиготности отца) вероятность образования резус-отрицательного плода с генотипом dd составляет 50%. Тогда никакой иммунизации не происходит. Вполне возможно, что и последующие дети окажутся резус-отрицательными, и никакого резус-конфликта не возникнет.
Во-вторых, даже если в браке ♀ dd × ♂ Dd первый плод окажется резус-положительный (Dd), и произойдет иммунизация материнского организма, то последующие дети могут оказаться резус-отрицательными, и резус-антигены не окажут на их эритроциты вредного воздействия.
В-третьих, при обнаружении в крови резус-отрицательной женщины резус-антигенов (например, сразу после рождения резус-положительного ребенка) ей переливают сыворотку крови от мужчин-добровольцев, искусственно иммунизированных резус-антигенами. Такая сыворотка содержит резус-антитела, которые разрушают эритроциты с резус-антигенами, появившиеся в женском организме. В результате иммунизации женского организма не происходит.
И, наконец, при рождении резус-положительного ребенка, в крови которого обнаруживаются резус-антитела, ему производят полное (заместительное) переливание крови на кровь, не содержащую резус-антитела.
2.4 Географическое распределение групп крови А1А2B0
Прежде всего, мы должны понять, что гены O, A, и B являются очень древними. Они не являются результатом недавних мутаций. Например, анализ египетских мумий показывает, что три гена, отвечающих за образование групп крови, присутствуют там в таких же пропорциях, как и у современных египтян.
На сегодня наиболее обоснованной является теория, согласно которой современный человек сформировался в Центральной Азии в то время, когда климатические условия там были более благоприятными, чем сейчас. Эта самая ранняя группа современных людей могла иметь гены с повторяемостью 25 A, 15 B, и 60 O на каждую сотню A-B-O генов групп крови. Примерно такова повторяемость генов в Центральной Азии сегодня.
По мере приближения к историческим временам и роста человеческой популяции на Земле, отток людей из Центральной Азии увеличивался. Эмиграционные группы были достаточно велики, чтобы перенести с собой ген B. Наиболее приближенные к Центральной Азии районы, например, Маньчжурия и северная Индия, получили наибольшую порцию. Восточная Европа получила большее количество на этом континенте, и по мере продвижения на запад частота гена B исчезает. Северо-восточная Африка получила больше всего на своем континенте, и частота гена B уменьшается по мере продвижения на запад и юг.
Народы Азии и Африки в общем имеют высшую частоту гена B, чем другие люди. Они отличаются друг от друга в частоте Rh. Азиаты имеют выше частоту гена, называемого Rhz, чтобы отличить его от других генов ряда Rh. С другой стороны, африканцы имеют высокую повторяемость другого гена Rh, называемого Rho.
Американские, австралийские и азиатские группы имеют небольшое количество или вообще не имеют гена rh. Африканская группа имеет небольшое количество гена rh. Жители Европы (включая американцев и австралийцев, ведущих свое происхождение от европейцев), тем не менее, имеют значительное количество гена rh; примерно каждый седьмой из них является Rh-отрицательным.
Если мутации, в результате которых появились антигены A и B являются древними, то группа крови O гораздо старше.
Другой параметр, свидетельствующий о древности группы O, следует из анализа физической антропологии и предполагает, что большую часть своего существования человечество принадлежало исключительно к группе O.
Новые исследования по митохондриальной DNA (mtDNA) подтверждают теорию о появлении Homo sapiens в Африке и последующем распространении в других регионах.
Высокий коэффициент группы крови O позволяет предполагать, что американские индейцы и эскимосы напрямую произошли от кроманьонских предков, вероятно, монгольских, которые мигрировали примерно в 15,000 до н.э. в Америку.
Малая часть коренных индейцев-американцев имеют группу крови В, таким образом они должно быть мигрировали в Америку достаточно поздно для получения положительного гена Rh, но слишком рано, чтобы получить ген для B.
Группа крови A в наибольших концентрациях обнаруживается у западноевропейцев. В отличие от групп крови B и O, группа A имеет множество вариаций. Основная группировка, A1, встречается примерно у 95 процентов группы A. Самая большая подгруппа, A2, встречается в основном на Северном Кавказе. A2 в очень больших концентрациях обнаруживается в Исландии и Скандинавии, особенно у лаппов, древнейших обитателей этого района. Они почти уникальны в высокой частоте группы A, и имеют самую высокую частоту A2, 42 процента в одной группе населения. A2 ген почти полностью сконцентрирован у населения Кавказа.
Частота группы А в Европе снижается по мере продвижения на восток. На большей территории Европы повторяемость гена А превышает 25 процентов. Он так же в значительных количествах обнаружен на территориях вокруг всего Средиземного моря, особенно на Корсике, Сардинии, Испании, Турции и Балканах. Ясно, что человечество чаще основывало постоянные поселения в тех местах, где условия обещали им самые высокие шансы на выживание.
Видимо ген В мог попасть на Индийский субконтинент через завоевания. При исследовании распределения ABO вдоль Шелкового пути Северо-западного Китая наблюдалось явное увеличение частоты группы крови В, особенно когда сравнивались субъекты монголоидний выборки с субъектами европеидной выборки.
Почти непрерывная цепь гористой территории протянулась от Уральских гор до Кавказа в Азии, и далее до Пиренеев в южной Франции. Этот барьер разбил миграцию групп крови на два основных маршрута; северный поток и южный. Переселенцы, выбравшие южный путь, стали предками жителей Средиземноморья и западноевропейцев, они несли с собой ген группы А. Уральские горы остановили большую миграцию на запад из Азии, хотя небольшое количество европеидов попало в восточную Европу, принеся с собой ген группы крови В, который они получили, смешавшись с азиатскими монголоидами. Это барьер послужил для разделения групп крови на западную A; и восточную B группы.
Монголоиды, носители группы крови B; продолжали двигаться на север, в направлении современной Сибири.
Для современных антропологов группа крови В продолжает и на сегодняшний день оставаться ‘Восточной’ группой крови. В больших количествах она обнаруживается у азиатского населения, такого как китайцы, индийцы, сибиряки. В Европе группа крови В чаще всего встречается у венгров, русских, поляков и других восточно-европейских народов. Среди до-неолитических народов, таких как баски и американские индейцы, группа крови В практически не встречается.
Из всех групп крови ABO, B демонстрирует четко выраженное географическое распространение. Протянувшись, как огромный пояс, вдоль равнин Евразии до Индийского субконтинента, группа крови В в повышенных количествах насчитывается на территориях от Японии, Монолии, Китая и Индии до Уральских гор. Отсюда в западном направлении процентное соотношение уменьшается, пока не достигает самого низкого значения в самой западной точке Европы.
Группа крови B является явно не индоевропейской группой крови. В Европе, только две области выделяются высоким соотношением группы крови В: одна среди не индоевропейских народов, известных как финно-угры (венгры и финны), другая – среди центрально-славянских народов (чехи, южные поляки, и северные сербы). Вторгающиеся Викинги также могли иметь относительно высокий процент гена В, поскольку многие города Британии и Западной Европы, связанные с побережьем только внутренними коммуникациями такими, как большие реки, имеют, в сравнении с окружающими территориями, непропорциональное количество группы В.
Небольшие количества групп крови у западноевропейцев отражают миграцию азиатских кочевников на запад. Это наиболее явно прослеживается у восточных западноевропейцев, немцев и австрийцев, которые имеют неожиданно высокий процент группы В по сравнению со своими западными соседями. Высокая повторяемость группы В у немцев имеет место на территории около верхней и средней Эльбы, важной естественной границы между ‘цивилизацией’ и ‘варварством’ в древние и средние века.
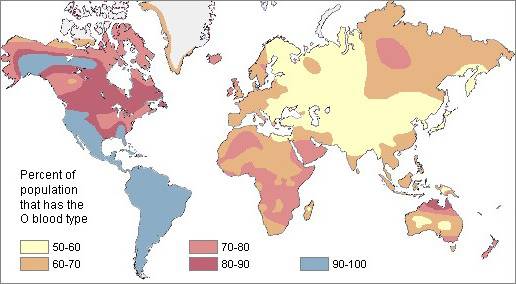
Рис. 1. Частота гена 0 групп крови системы АВ0
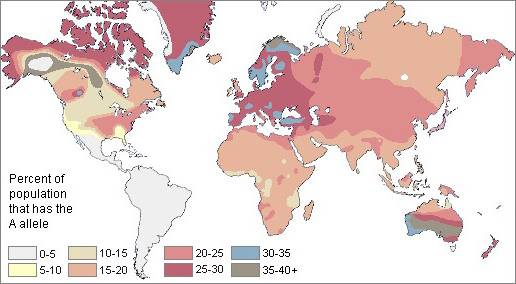
Рис. 2. Частота гена А групп крови системы АВ0
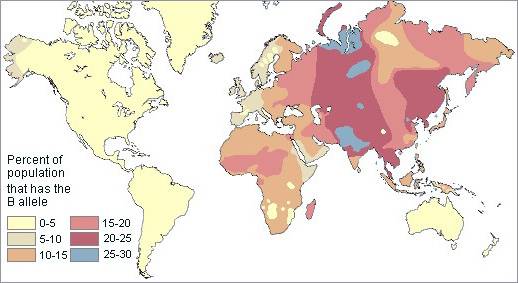
Рис. 3. Частота гена В групп крови системы АВ0
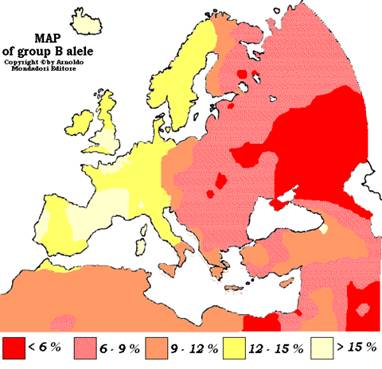
Рис. 4. Распределение аллеля В у населения Европы
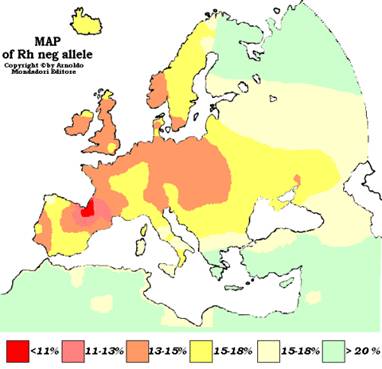
Рис. 5. Распределение групп крови по системе Rh у населения Европы
кровь группа полиморфизм аллель
3. ИМУНОХИМИЧЕСКИЕ ХАРАКТЕРИСТИКИ ГРУПП КРОВИ
Принадлежность индивидуума к той или иной группе крови определяется наличием или отсутствием в эритроцитах группоспецифических веществ – антигенов А и В. Вещества с такими свойствами и такой же специфичностью были найдены в большинстве тканей организма. Исключения составляют яички, хрусталик, хорион, плацента, хрящ и эпителий кожи. Группоспецифические вещества тканей относятся к так называемым спирторастворимым и представляют собой гликолипиды. Водорастворимые группоспецифические соединения в больших количествах встречаются в различных слизистых секретах (слюне, слизи желудочно-кишечного тракта и др.) и являются гликопротеинами. Важно подчеркнуть, что антигены А и В обнаруживаются в семенной жидкости, плазме, слюне и женском молоке, причем именно в жидкостях выявляется в растворенном состоянии наибольшее количество этих веществ. Таким образом, спирто- и водорастворимые субстанции относятся к различным классам макромолекул, обладающих одинаковыми антигенными свойствами, детерминированными одним и тем же локусом.
Химическая структура определена для водорастворимых группоспецифических веществ, которые состоят из большого числа относительно коротких олигосахаридных цепей, соединенных ковалентными связями с полипептидными основами. Молекулярная масса этих гликопротеидов – около 500 000 дальтон, их углеводные цепи состоят из 7…8 остатков сахара, и каждая молекула содержит около 300 углеводных цепей. Групповую специфичность антигенов А и В определяют терминальное положение сахаров в цепи молекулы, причем существенно, что молекула вещества А заканчивается остатком N-ацетилгалактозамина, а цепь молекулы антигена В остатком галактозы. В остальном обе цепи идентичны. При 0-группе крови в цепи отсутствуют терминальные остатки, т. е. N-ацетилгалактозамин или галактоза, и молекула приобретает так называемую Н-специфичность. Установлено, что у носителей аллеля IА имеется специфическая трансфераза, переносящая N-ацетилгалактозамин к концевой группе олигосахаридной цепи. Аналогичным образом у носителей аллеля IВ, обладающих D-галактозилтрансферазой, к концевой группе олигосахаридной цепи присоединяется галактоза. У индивидуумов с группой крови 0, т. с. гомозигот по аллелю I0 соответствующие трансферазы отсутствуют, и к углеводной цепи никакой дополнительный остаток не присоединяют. Без этого дополнительного остатка иммунологически определяется Н-антигенная специфичность. Н-антигены как продукты промежуточной реакции присутствуют у индивидуумов с А- и В-группами крови, но естественно, что концентрация Н-антигенов выше