


Влияние повышенного и сниженного уровня моноаминов на функциональную организацию колонок C1 коры мозга крысы
В.Н. Ласков
НИИ нейрокибернетики им. А.Б.Когана, РГУ, Россия
Повышение внутримозгового содержания нейромодулятора серотонина после электрической стимуляции ядер шва, а также снижение содержания биогенных моноаминов после внутрибрюшинного введения резерпина вызывали разнонаправленную пластификацию синаптических связей во входных, выходных и ассоциативных нейронных ансамблях бочонковой колонки С1 коры мозга крысы и модулировали уровень ее возбуждения за счет изменения количества фоновоактивных нейронов, а также частоты и структуры их импульсных разрядов. Показана зависимость характера сдвигов частоты, структуры импульсных разрядов, плотности кросскорреляционной взаимосвязи соседних нейронов от предыдущего уровня возбуждения колонки. Новизна работы состоит в модульном аспекте рассмотрения моноаминергических механизмов синаптической пластичности корковых нейронов в контексте действующего в пределах фокальной бочонковой колонки гипотетического авторитмичного сканирующего механизма формирования временной связи.
Механизмы регуляции уровня возбуждения корковых нейронов и модуляции эффективности их связей опосредованы наряду с холинергическими нейромедиаторными системами также и влияниями со стороны нейромодуляторных структур ствола мозга, синтезирующих моноамины: серотонин (СТ), норадреналин и дофамин /2,4,5,6,12,23/. СТ, который транспортируется к коре мозга из ядер шва, рассматривается в качестве нейромодулятора, обеспечивающего механизмы положительного подкрепления посредством стабильного повышения при обучении уровня возбуждения и кросскорреляционного взаимодействия корковых нейронов /4,16,18/. Методами электронной микроскопии показано, что терминали проецирующихся к неокортексу восходящих СТ-ергических пучков не образуют типичных синаптических контактов на мембране нейронов. Они в виде варикозных расширений на большом протяжении выстилают двойным слоем плотно заполненное перикарионами, дендритами и глией межнейронное пространство, и обеспечивают непосредственный контакт транспортируемого моноамина с синапсами специфических и неспецифических афферентов /14,15/. Возбуждение СТ-ядер вызывает быстрое насыщение пресинапсов, которые не имеют собственной постсинаптической комплиментарной части.
Синапсы такой химической природы и конструкции больше не встречаются ни среди корково-корковых, ни среди окончаний иных проекционных систем мозга. Это свидетельствует об их особой роли в обеспечении мозговых функций. Высказано предположение, что моноаминергические системы специфично и адресно модулируют эффективность синапсов первичных специфических, неспецифических и ассоциативных афферентов. Тем самым они обеспечивают необходимый уровень пластичности межнейронных взаимодействий при формировании временных связей и памятных следов /4,16,18/. Модулирующие влияния восходящей моноаминергической системы опосредованы через регуляцию уровня возбуждения корковых нейронов за счет активации вторичных посредников - циклических нуклеотидов /17/, что обеспечивает стабилизацию пластифицированных в ходе обучения межнейронных связей.
Плодотворной для развития идеи о модулировании синаптической эффективности со стороны моноаминергических структур оказались математическая и нейрофизиологическая модели Жадина М.Н. /4,5/, описывающие динамику активности корковых нейронов при длительном воздействии положительного и отрицательного подкрепления на нейроны коры мозга. В основу модели положена гипотеза /4,16,18/, согласно которой СТ- и норадреналинергическая системы являются конечными звеньями, соответственно, положительного и отрицательного подкрепления, вызывающего устойчивые изменения эффективности корково-корковых синапсов. В модели при длительном действии моноамина, обеспечивающего положительное подкрепление, любые превышающие средний уровень изменения уровня активности нейронов доводятся до своих крайних проявлений - предельно высокому и предельно низкому. Баланс возбуждения и торможения при такого рода положительном подкреплении характеризуется крайней неустойчивостью: под влиянием каждой достаточно мощной афферентной посылки в системе происходят быстрые переходы от одного уровня возбуждения на другой. В присутствии моноамина, обеспечивающего отрицательное подкрепление, все происходит с точностью до наоборот: и сильно, и слабо возбуждающиеся нейроны в соответствии с предложенной схемой переходят на устойчивый к разного рода афферентным посылкам промежуточный уровень возбуждения.
В соответствии с нейрофизиологическими данными поведение корковых нейронов в присутствии СТ и норадреналина согласуется с предложенной математической моделью /2,3,5,7/. В работе /12/ длительная аппликация СТ и норадреналина вызывала более сложные по характеру изменения структуры фоновой импульсной активности нейронов зрительной и сенсомоторной областей коры мозга у кроликов, чем простое возбуждение и торможение в ответ на аппликацию нейромедиаторов ацетилхолина и ГАМК. Были также продемонстрированы различные типы реакций корковых нейронов на один и тот же нейромодуляторный моноаминергический стимул: конечный результат при этом обнаружил зависимость, как и предсказано моделью, от исходного уровня возбуждения коркового нейрона. Тем самым подтверждены представления о том, что нейромодуляторы в отличие от нейромедиаторов, действие которых локально и кратковременно, не вызывают простого возбудительного или тормозного эффектов, а влияют на текущую деятельность нейрона, изменяя ее в течение длительного времени в том или ином направлении /1,20,21/.
Краткий обзор наиболее значимых работ, посвященных исследованию механизмов модулирования синаптической эффективности в реальных нейронных сетях и их моделях, свидетельствует о важной роли моноаминергических систем мозга в нейрохимическом обеспечении его ассоциативных функций. Вместе с тем многие аспекты роли и механизмов участия моноаминергических систем в условнорефлекторной деятельности, а также в процессах самоорганизации элементарных нейронных ансамблей /8/ остаются не ясными, либо требуют уточнения. В данной работе исследованы нейронные корреляты временной связи при пониженном после внутрибрюшинного введения резерпина и повышенном после электрической стимуляции ядер шва уровне возбуждения фокальной колонки представительства условного стимула (УС) в С1 коры мозга крыс.
Методика
Опыты выполнены на белых беспородных крысах обоего пола весом 120-200 г, оперированных под эфирным наркозом, обездвиженных тубокурарином (1,5 мг/кг в/м) и переведенных на искусственное дыхание. Трахеотомия не проводилась: воздух в легкие нагнетался через ноздри. Использована местная анестезия (0,5% р-р новокаина) операционного поля. Голова крысы фиксировалась с помощью игольчатых головодержателей. Череп трепанировался над постеромедиальной субзоной бочонков (ПМСБ) корковой области С1 по координатам: 2-3 мм каудальнее брегмы и 5-6 мм латеральнее сагиттального шва. Фоновую импульсную активность нейронов и фокальные ВП регистрировали одиночными стеклянными микроэлектродами (электролит - 2,5 М р-р NaCl) с сопротивлением не ниже 5 мОм, а в ряде случаев склеенными по три-пять вольфрамовыми в стеклянной изоляции микроэлектродами (0,5-1 мОм, расстояние между кончиками - 70-100 мкм), которые погружались в кору перпендикулярно ее поверхности с помощью наклонного манипулятора, оснащенного шаговым двигателем. Регистрацию осуществляли при полосе 2-2000 Гц на входных фильтрах электроэнцефалографа 4ЭЭГ-03. Импульсная активность после предварительного формирования на 4-канальном амплитудном анализаторе АА-4 записывалась на 14-канальный магнитограф EАМ-500 (ЧССР, г. Прага) и вводилась в персональный компьютер IBM-PC/AT-386 для текущей и последующей обработки.
В качестве адекватного стимула использовано сгибание центральной в РП вибриссы с удержанием ее в отклоненном положении в течение 0,1-1,0 с. Чтобы избежать неконтролируемых смещений, дальние от носа вибриссы, достигающие в длину 30-50 мм, билатерально укорачивались до 5-10 мм. Механический датчик представлял собой щуп, приклеенный на свободном конце пьезокерамической пластины, изготовленной в ОКБ "Пьезоприбор" РГУ, на которую подавалось напряжение от 60 до 1 В с выхода ЭСЛ-2, задающего амплитуду отклонения на конце щупа от 90 до 1,5 мкм. Метрологическая проверка показала линейную зависимость напряжения и амплитуды сгибания пьезокерамической пластины в используемом диапазоне, поэтому в опыте амплитуда отклонения щупа оценивалась по напряжению на выходе ЭСЛ-2.
Контролем попадания электродов в ПМСБ служили фокальные ВП, возникающие в ответ на легкое постукивание по контралатеральным вибриссам вручную, а затем с помощью тактильного датчика, запускаемого от ЭСЛ-2. Идентификация бочонковых колонок под каждым из микроэлектродов производилась следующим образом. Сравнивали наблюдаемые на мониторе фокальные ВП по их латентным периодам (ЛП), крутизне и амплитуде в ответ на последовательное сгибание вибрисс и устанавливали соответствие между корковым бочонком и центральной в РП вибриссой. Для уточнения идентификации центра РП производилась стимуляция ближайших к центру вибрисс со ступенчатым уменьшением амплитуды их отклонения. Серия стимулов определенной амплитуды предъявлялась ритмично с частотой 0,5 Гц. По 10-20 реализациям строились точечные или столбиковые перистимульные гистограммы (ПСГ) с бином 1-2 мс за период 100-500 мс. Сужение РП до одной вибриссы в случае ее центрального положения имело место при более низких амплитудах ее отклонения - пороговый тест /9/.
Определяли удельное количество фоновоактивных нейронов, по которому сравнивали исходный уровень возбуждения бочонковой колонки с уровнем, устанавливающимся после стимуляции ядер шва и в последействии резерпина. Нейроны с фоновой активностью исследованы в параллельных проходках, которые осуществлялись через всю толщину коры при межэлектродном расстоянии в склейке - 100 мкм. Треки располагались во фронтальных (AP - 2,0 и AP - 3,0) и сагиттальных (L - 5,0 и L - 6,0) плоскостях. На каждом из треков контролировали 60 точек с интервалом по глубине 30 мкм. Через 2-3 ч после введения резерпина (в/б 2,5 мг/кг), когда в соответствии с многочисленными литературными источниками наблюдается наиболее значительное снижение в мозге уровня СТ, и через 10 и 20 мин после электрической стимуляции ядер шва (частота 1 Гц, длительность серии стимулов 20 с, импульсы длительностью 0,1-1,0 мс, напряжение на выходе стимулятора 15 В) в тех же треках проводилась повторная регистрация фоновых импульсных разрядов. В ряде случаев была уверенность, что тот же нейрон наблюдали до и после введения препарата. В каждом опыте исследовалась также непосредственная динамика импульсной активности наиболее многочисленной группы нейронов, регистрируемых с помощью микроэлектродной склейки одновременно.
Координаты ядер шва определяли по атласу /19/. Стимулирующие электроды погружались с помощью микроманипулятора ММ-1 и под контролем бинокулярного микроскопа МБС-2 по координатам: АР - 6,6; L - 0,4; H - 4,5, с контролем погружения по индикатору глубины ГИ-100. Твердая мозговая оболочка предварительно удалялась. Стимулирующие монополярные полумикроэлектроды представляли собой вольфрамовые в стеклянной изоляции электролитически заточенные проволоки диаметром 50 мкм с сопротивлением 0,5 мОм. Верификацию треков стимулирующих электродов в ядрах шва осуществляли морфологически на парафиновых и замороженных с помощью сухого льда срезах. Выработка условного мигательного рефлекса (УР) осуществлялась по схеме классического обусловливания. Длительность УС и безусловного стимула (БС) равнялась 1 с, интервал между ними - 2 с, а сочетания УС и БС подавались с частотой 0,1 Гц. По каждому десятку стимулов строились ПСГ с квантом 2 мс за эпоху анализа 100 мс или 200 мс в фоне и 900 мс или 800 мс, соответственно, после предъявления УС. Реакции в ответ на обдувание роговицы при выработке мигательного УР, как правило, не регистрировались. Уровень выработки УР ограничивали предъявлением 50-60 подкреплений.
Частоту фоновой импульсации и характер взаимосвязи соседних корковых нейронов методом перекрестных интервальных гистограмм (ПИГ) исследовали до и в разные сроки после электрической стимуляции ядер шва. Частота импульсации нейронов измерялась с помощью частотомера Ч3-36 за 5-секундные интервалы времени.
Запуск стимуляции, амплитудная дискриминация, формирование и запись импульсной активности, а также оцифровка, построение и распечатка ПСГ, гистограмм межимпульсных интервалов (ГМИ) и ПИГ осуществлялись программными средствами. Статистическая обработка ПСГ состояла в вычислении с помощью программы "Statgraf" средней частоты импульсации, стандартной ошибки средней и достоверности различия средних по критерию Стьюдента: между фоновой и вызванной активностью - для установления эффективности каждого из стимулов; между реакциями по ходу выработки мигательного УР - для выявления условнорефлекторной пластичности. Пики на гистограммах считали значимыми, если они удовлетворяли критерию 2-сигма по отношению к среднему значению количества спайков в бине. Достоверность различий между выборочными распределениями оценивали по критерию Фишера.
Результаты исследования
Влияние электрической стимуляции ядер шва.
С помощью метода пошагового определения плотности активных в фоне нейронов в отвесных проходках через кору мозга исследованы в трех острых опытах 15 микроэлектродных треков, которые проходили, как показала пороговая идентификация, в пределах бочонковых колонок представительства вибрисс рядов В, С и D.
В первом опыте в контроле наблюдалось 39, во втором опыте - 37 и в третьем - 35 фоновых нейронов, что составило в контроле в среднем на один трек около 7 нейронов. Более плотно активные в фоне нейроны располагались на глубине 600-1500 мкм - 78% от общего количества зарегистрированных в контроле нейронов находились на этом уровне. Суммарно по трем опытам в контроле было зарегистрировано на уровнях: 2-3 слоев - 13 нейронов, 4 слоя -23 нейрона, 5 слоя - 52 нейрона, 6 слоя - 23 нейрона. В исходном состоянии на треках встречалось от 2 до 13 фоновоактивных нейронов. Как правило, треки с высокой плотностью нейронов были разделены одним-двумя относительно "пустыми" треками, принадлежавшими одноименной бочонковой колонке. В пределах одноименной колонки за одну проходку контролировали импульсную активность на 2-3 соседних треках. В девяти треках с высокой плотностью элементов (от 7 до 13) зарегистрировано 90 фоновых нейронов (82%), а в шести "пустых" (от 2 до 5 нейронов) - 21 нейрон (18%).
Электрическая стимуляция ядер шва приводила к возникновению у части молчащих нейронов спонтанных разрядов в течение 1-2 мин, а в ряде случаев до 20 мин и более. После электрической стимуляции на тех же треках в первом опыте зарегистрирована активность 43, во втором опыте - 40 и в третьем - 38 нейронов, что составило в среднем около 8 нейронов на трек. Из них зарегистрировано на уровнях: 2-3 слоев - 25 нейронов, 4 слоя - 24 нейрона, 5 слоя - 49 нейронов, 6 слоя - 23 нейрона. В 9 треках с высокой плотностью элементов при этом зарегистрировано - 87 нейронов (72%), а в "пустых" треках - 34 нейрона (28%). Из данных послойного и потрекового анализа следует, что электрическая стимуляция ядер шва наиболее значительно увеличивает количество фоновоактивных нейронов в бочонковых колонках на уровне 2-3 слоев - в 2 раза, и в "пустых" треках - в 1,6 раза.
Анализ динамики частоты фоновой импульсной активности корковых нейронов в контроле и после активации шва, результаты которого представлены в таблице, выявил послойную неравномерность в росте уровня возбуждения колонок. В наибольшей степени частота фоновой импульсации увеличивалась у выходных нейронов 5 - 6 слоев (P < 0,001), тогда как у ассоциативных нейронов 2-3 слоев повышение частоты было достоверным при более низком уровне значимости (P < 0,05). У нейронов афферентного уровня снижение средней частоты фонового разряда было недостоверным.
Таблица
Изменение частоты фоновой импульсной активности нейронов разных уровней бочонковой колонки С1 коры мозга крыс при электрической стимуляции ядер шва и под влиянием резерпина.
| Уровни | Количество нейронов | Частота (имп/с) | Количество нейронов | Частота (имп/с) |
| колонки | До стимуляции (контроль) | После стимуляции (опыт) | ||
| 2-3 слои | 16 | 13,7 | 16 | 17,6 |
| 4 слой | 25 | 9,7 | 20 | 9,1 |
| 5-6 слои | 38 | 12,4 | 25 | 20,9 |
| N = 79 | N = 61 | |||
| До введения резерпина (контроль) | После введения резерпина (опыт) | |||
| 2-4 слои | 33 | 11,4 | 28 | 8,4 |
| 5-6 слои | 48 | 9,5 | 36 | 7,4 |
| Суммарно | 81 | 9,4 | 64 | 7,2 |
Примечание: * - различия опыт-контроль достоверны при P< 0,05,
** - различия опыт-контроль достоверны при P < 0,001.
О перестройках структуры фонового импульсного разряда корковых нейронов при активации ядер шва свидетельствуют данные анализа ГМИ. На рис. 1 представлены в качестве типичных примеров ГМИ импульсации трех нейронов, соответственно, с верхнего - ассоциативного, среднего - афферентного и нижнего - эфферентного уровней бочонковой колонки С3 в контроле, а также через одну, две и три минуты после электрической стимуляции ядер шва.
У ассоциативных нейронов на уровне 2-3 слоев (рис.1,А) перестройки структуры фоновых импульсных разрядов были, как правило, динамичными, протекали волнообразно с возвратом к контрольному аритмичному паттерну. Так, у нейронов 2-3 слоев (N=10) средний межимпульсный интервал был равен в контроле 15,9 ± 3,0 мс, через минуту после стимуляции ядер шва возрастал до 21,7± 4,1 мс (P<0,05), через две минуты возвращался к контрольному уровню - 16,4± 2,7 мс, а через три минуты снова достоверно (P<0,05) увеличивался до 27,6± 3,2 мс.
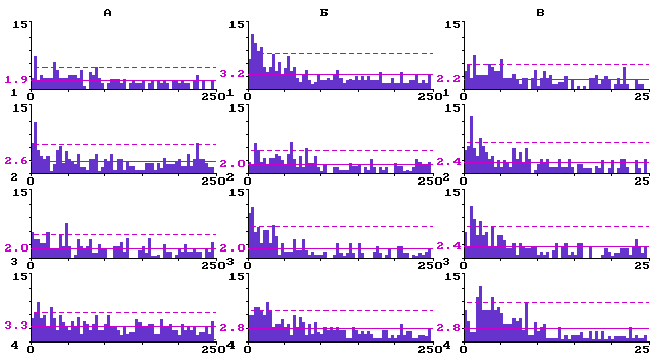
Рис.1. Перестройка структуры фоновой импульсной активности нейронов разных уровней бочонковой колонки С1 коры мозга крыс при электрической стимуляции ядер шва.
Гистограммы межимпульсных интервалов: А - для нейрона ассоциативных слоев; Б - для нейрона афферентного слоя; В - для нейрона эфферентного слоя; 1 - до стимуляции ядер шва; 2 - через одну минуту; 3 - через две минуты; 4 - через три минуты после стимуляции. По вертикали - количество интервалов; по горизонтали - их длительность в мс. Сплошная горизонтальная линия - среднее количество интервалов; штриховая - уровень 2s .
У входных фоновоактивных нейронов 4 слоя (рис.1,Б) исходные пачечные и группированные импульсные разряды становились единично-аритмичными. Средний межимпульсный интервал у входных нейронов (N=10) в контроле был равен 26,7± 5.2 мс. Через одну минуту после стимуляции ядер шва он достоверно (Р<0,05) сокращался до 16,4± 3,4 мс, через две минуты удерживался на том же уровне - 16,4± 5,0 мс, а через три минуты был равен в среднем 23,0± 4,1 мс, что по t- критерию недостоверно (Р<0,05) отличается от контроля. Оценка достоверности различия выборочных распределений межимпульсных интервалов для входных нейронов по критерию Фишера показала достоверную (P<0,05) трансформацию структуры их фонового импульсного разряда через одну минуту после стимуляции ядер шва (F=2,36 при Fst=1,39), возврат к контрольному паттерну через две минуты (F=1,07 при Fst=1,39) и повторную рандомизацию разрядов к третьей минуте (F=1,58 при Fst=1,39).
У выходных нейронов слоя 5 (рис.1,В) наблюдалась прямо противоположная тенденция: их исходный единичный аритмичный разряд после электрической стимуляции ядер шва перемежался вначале пачками (через одну и две минуты), а к третьей минуте - группами фоновых импульсов. Средний межимпульсный интервал у выходных нейронов (N=12) в контроле был равен 18,3± 3,6 мс, через одну минуту после стимуляции ядер шва - 20,2± 4,8 мс, через две минуты - 19,7± 4,7 мс, через три минуты - 23,5± 6,0 мс, что по t- критерию во всех трех случаях недостоверно отличается от контроля (P<0,05). Оценка достоверности различия выборочных распределений межимпульсных интервалов для выходных нейронов по критерию Фишера показала однонаправленную, в течение трех наблюдаемых минут, достоверную (P<0,05) трансформацию структуры единичного аритмичного их разряда в пачечный и групповой (F=1,70; 1,65; 2,66, соответственно, после первой, второй и третьей минуты при Fst=1,39). Такого рода перестройки структуры фоновых разрядов в исследованных выборках входных и выходных нейронов были стабильными в течение десятков минут.
В выборке попарно зарегистрированных нейронов (N=28) оценивались влияния активации ядер шва на характер межнейронных взаимодействий с использованием метода построения ПИГ. Значимые пики на ПИГ рассматривали как проявление статистически выявленной функциональной связи между нейронами, осуществляющейся с определенной задержкой и в определенном направлении.
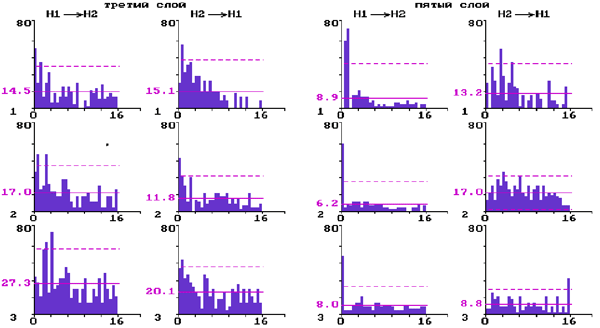
Рис.2. Перестройки межнейронных взаимодействий нейронов на разных уровнях бочонковой колонки С1 коры мозга крыс при электрической стимуляции ядер шва.
Постимпульсные гистограммы связи для прямых и обратных влияний парных нейронов слоя 3 и парных нейронов слоя 5:
1 - до стимуляции ядер шва; 2 - через десять минут; 3 - через двадцать минут после стимуляции. По вертикали - количество импульсов; по горизонтали - время в мс. Сплошная горизонтальная линия - среднее количество импульсов; штриховая - уровень 2s .
Для верхних слоев показано (рис. 2) появление на ПИГ после электрической стимуляции дополнительных длиннолатентных значимых пиков и уменьшение пиков с нулевой латентностью. В нижних слоях при этом наблюдалось ослабление как прямых, так и обратных межнейронных моно- и полисинаптических влияний и появление односторонних влияний с нулевой и высокой латентностью.
Сниженное под влиянием резерпина внутримозговое содержание СТ.
Анализ средней текущей частоты фоновой импульсной активности нейронов бочонковой колонки С1 до и после введения резерпина, результаты которого представлены в таблице, показал, что на пике действия резерпина суммарно по колонке, и на каждом из ее уровней имеет место достоверное (Р<0,05) снижение частоты импульсации. При этом характерный для контроля более высокий частотный уровень нейронов поверхностных слоев по сравнению с глубокими слоями на фоне резерпина выравнивался. Следует отметить, что около 18 % нейронов, исходная частота импульсации которых не превышала средних значений по выборке (Р<0,05), были резистентными к резерпину. У 10 % нейронов, которые зарегистрированы преимущественно в эфферентных слоях, частота импульсации в последействии резерпина достоверно (Р<0,05) повышалась. Их исходная частота превышала среднее значение по всей выборке (Р<0,01).
На рис. 3 представлены ГМИ фоновой импульсной активности трех нейронов, принадлежащих бочонковой колонке С3. Нейрон на (А) имел до введения резерпина три отчетливых максимума в ГМИ, что свидетельствует о наличии в структуре его исходного разряда пачек и групп, перемежающих аритмично следующие единичные разряды. Через 2,5 ч после введения резерпина частота импульсации
данного нейрона уменьшилась, а структура рандомизировалась - исчезли пачечные и групповые разряды. Средний межимпульсный интервал у снижающих частоту нейронов (N=10) в контроле был равен 19,8± 4,4 мс, а через 2-3 часа после введения резерпина - 24,5± 2,6 мс, что по t- критерию недостоверно отличается от контроля (P<0,05).
У нейрона на (Б) через 2 ч после введения резерпина ГМИ стала островершинной с двумя достоверными максимумами в диапазоне самых малых интервалов, что отражает появление в структуре непрерывного аритмичного разряда коротких пачек и разной длительности групп импульсов. Такая динамика структуры фонового разряда была характерна для нейронов, которые увеличивали под влиянием резерпина частоту импульсной активности. Средний межимпульсный интервал у повышающих частоту нейронов (N=10) в контроле был равен 22,4± 6,6 мс, а через 2-3 часа после введения резерпина - 19,6± 3,9 мс, что по t- критерию недостоверно отличается от контроля (P<0,05).
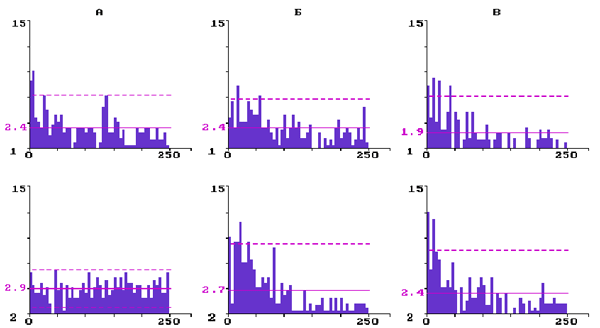
Рис.3. Перестройка структуры фоновой импульсной активности трех нейронов разных уровней бочонковой колонки С1 коры мозга крыс при действии резерпина
Гистограммы межимпульсных интервалов: А - при уменьшении частоты импульсации; Б - при увеличении частоты; В - при стабильной частоте; 1 - в контроле; 2 - в последействии резерпина. По вертикали - количество интервалов; по горизонтали - их длительность в мс. Сплошная горизонтальная линия - среднее количество интервалов; штриховая - уровень 2s .
На (В) представлен нейрон, у которого в течение 3 ч после введения резерпина частота фонового разряда существенно не менялись, а его структура изменялась недостаточно отчетливо. Резистентные к резерпину нейроны преимущественно залегали на уровне 4-го слоя колонки, т.е. они в большинстве своем были входными для первичной специфической афферентации. Средний межимпульсный интервал у резистентных к резерпину нейронов (N=10) в контроле был равен 15,4± 5,0 мс, а через 2-3 часа после введения резерпина - 20,2± 6,0 мс, что по t- критерию недостоверно отличается от контроля (P<0,05).
Оценка достоверности различия выборочных распределений межимпульсных интервалов у представленных трех типов нейронов по критерию Фишера показала достоверную (P<0,05) трансформацию под влиянием резерпина структуры их фонового импульсного разряда (F=1,76; 2,88; 1,46, соответственно, для А, Б и В при Fst=1,45).
С помощью метода пошагового определения плотности активных в фоне нейронов в отвесных проходках через кору мозга исследованы в трех острых опытах 9 микроэлектродных треков, которые проходили, как показала пороговая идентификация, в пределах бочонковых колонок представительства средних вибрисс рядов В и С. В первом опыте в контроле наблюдалось 29, во втором опыте - 27 и в третьем - 25 фоновых нейронов, что составило в контроле в среднем на один трек 9 нейронов. Более плотно активные в фоне нейроны располагались на глубине 500-1500 мкм - 70% от общего количества. Суммарно по трем опытам в контроле было зарегистрировано на уровне: 2-3 слоев - 18 нейронов, 4 слоя -26 нейронов, 5 слоя - 23 нейрона, 6 слоя - 14 нейронов. В исходном состоянии на треках встречалось от 4 до 18 фоновоактивных нейронов. Как и в серии опытов с электрической стимуляцией ядер шва, треки с высокой плотностью нейронов были разделены относительно пустыми" треками. В шести треках с высокой плотностью элементов (7 - 18) зарегистрировано 67 фоновых нейронов (83%), а в трех относительно "пустых" треках (4 - 5 нейронов) - 14 нейронов (17%).
Пространственно регулярный профиль распределения фоновоактивных нейронов в ПМСБ CI коры мозга крыс сохранялся и на фоне резерпина. Спустя 2-3 часа после введения резерпина в тех же треках в первом опыте зарегистрирована активность 21 нейрона, во втором опыте - 22 нейронов и в третьем - 21 нейрона, что составило в среднем около 7 нейронов на трек. Из них зарегистрировано на уровне: 2-3 слоев - 10 нейронов, 4 слоя - 23 нейрона, 5 слоя - 21 нейрон, 6 слоя - 10 нейронов. В шести треках с высокой плотностью элементов при этом зарегистрировано - 52 нейрона (81%), а в трех "пустых" треках - 12 нейронов (19%). Из данных послойного анализа следует, что резерпин наиболее существенно уменьшает количество фоновоактивных нейронов в бочонковых колонках на уровне 2-3 слоев (на 44%) и 6 слоя (на 29%). Плотные и относительно "пустые" треки в последействии резерпина содержали, соответственно, на 27% и на 25% меньше активных в фоне нейронов.
Структура фазических возбудительных реакций на отклонение центральной в РП вибриссы на фоне резерпина характеризовалась у 63 % нейронов выборки меньшей по сравнению с контролем частотой и большей длительностью первичного и вторичного возбуждения, а также разделяющей их тормозной паузы. Наряду с этим у многих нейронов с высоким уровнем фоновой импульсной активности (57 %) частота вызванной импульсной активности после введения резерпина достоверно (P < 0,05) увеличивалась, а средняя длительность фаз первичного и вторичного возбуждения и разделяющей их тормозной паузы сокращалась.
На рис. 4 графиками представлены распределения значений тормозной паузы в фазических возбудительных реакциях одного из нейронов (колонка С3, на уровне слоя 5) по частоте их реализуемости при повторном изолированном и сочетаемой с подкреплением отклонении центральной в РП вибриссы. Показано, что в контроле до введения резерпина (А) при изолированном предъявлении УС не обнаруживается отчетливой зависимости между длительностью тормозной паузы и частотой ее реализуемости в серии повторяющихся стимулов. Однако при выработке мигательного УР в распределении появляется максимум в диапазоне (140-180) мс, в котором находится около 40 % всех значений. Распределение значений тормозной паузы в реакциях этого же нейрона на изолированное предъявление УС в последействии резерпина (Б) апроксимируется также наклонной прямой линией, но возрастающей в направлении минимальных значений исследуемого параметра.
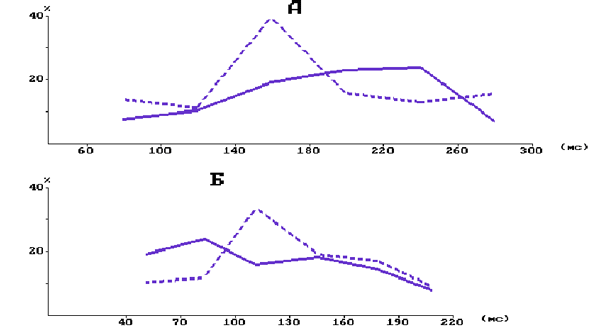
Рис. 4. Распределение значений тормозной паузы в фазированных возбудительных реакциях на УС нейронов фокальной бочонковой колонки С1 коры мозга крысы.
А - при выработке мигательного УР в контроле; В - в последействии резерпина. Сплошная линия - при изолированной тактильной стимуляции; пунктирная - при подкрепляемом предъявлении УС. По вертикали - количество значений длительности тормозной паузы в %; по горизонтали - длительность тормозной паузы в мс.
При выработке мигательного УР на фоне резерпина в распределении значений тормозной паузы, более сглаженный по сравнению с контрольным максимум, возникает в другом диапазоне - (100-130)мс. У данного нейрона под влиянием резерпина размах варьирования значений тормозной паузы становился более узким - (40-220) мс по сравнению с контролем (60-300) мс. Средняя длительность тормозной паузы по всей выборке в контроле на изолированное предъявление УС равнялась (147,7± 0,7) мс, а на подкрепляемые обдуванием роговицы была выше (P < 0,001) - (182,7± 0,4) мс. Средняя длительность тормозной паузы в последействии резерпина при изолированной стимуляции была выше (P < 0,05) по сравнению с контролем - (169,7± 1,1)мс, а при подкрепляемой обдуванием роговицы - не отличалась ни от контрольного значения, ни от значения при неподкреплении УС и равнялась (175,7± 1,0)мс.
Обсуждение результатов
Уникальные гистологические особенности ПМСБ представительства вибрисс в С1 коры мозга многих грызунов и других млекопитающих вызывают повышенный интерес к функциональным свойствам проекционных модулей данной области. Относительно простая их идентифицируемость на порядок повышает точность всех аспектов нейрофизиологического анализа. В предыдущих наших работах /10,11,22/ изучались локальные механизмы замыкания временной связи в фокусе коркового представительства условного стимула, который был ограничен отдельной бочонковой колонкой в С1 мозга крыс. Была показана структура межнейронных, вертикальных и горизонтальных, внутриколонковых и межколонковых взаимодействий в динамике выработки мигательного УР, определены зависимости скорости и характера динамики, а также степени упрочения временной связи от разнообразных форм пластичности паттернов импульсной активности нейронов, имеющих разную топологию в бочонковой колонке.
На основании данных о динамике и взаимной обусловленности отдельных компонентов фазических нейронных реакций фокуса коркового представительства УС на разных этапах обучения была сформулирована гипотеза о механизме авторитмичного сканирующего ассоциирования фокальной колонки с другими колонками, имеющими отношение к обработке параметров сочетаемых при обучении стимулов /10,11,22/. Условнорефлекторная перестройка структуры межнейронных связей в пределах фокальной корковой колонки, согласно такому предположению, состоит в ограничении ее авторитмичных сканирующих взаимодействий в распределенной системе заинтересованных структур. Это приводит к повышению вероятности конвергенции и интеграции синфазных волн в