


Биологические основы выращивания белуги (Huso huso)
Аквакультура в мире в начале XXI века характеризуется прогрессирующим ростом. Это происходит на фоне резкого падения запасов естественных популяций рыб в связи с их сверхинтенсивным промышленным и браконьерским ловом, разделом Каспия на отдельные территории суверенных государств.
С введением новых правил рыболовства на Каспии с 1962 г. промысел рыбы был полностью перебазирован из прибрежья моря в реки. При этом в западно-каспийском районе, в том числе и в реках, промысел осетровых был полностью прекращен. Эта мера сыграла исключительно важную роль в сохранении молоди ценных промысловых рыб, особенно осетровых, восстановлению их запасов, в существенной мере подорванных к этому времени.
Концепцией развития рыбного хозяйства на Каспии, принятой в 50-е годы XX века, было предусмотрено развитие отрасли по трем основным направлениям:
Главное направление — пастбищное осетровое хозяйство — в основном должно было базироваться на искусственном разведении молоди на специализированных осетровых рыбоводных заводах и выпуске ее в море на нагул.
Второе направление — это осуществление комплекса рыбоводно-мелиоративных мероприятий по поддержанию запасов полупроходных видов рыб, размножающихся в дельтах рек.
Третье направление – развитие пресноводной аквакультуры на основе прудового рыбоводства.
Главной проблемой каспийского рыбного хозяйства в настоящее время является критическое состояние запасов осетровых, вызванное масштабным незаконным выловом во всех регионах моря.
По данным правительства Р.Ф. основными целями и задачами развития аквакультуры:
- усовершенствовать и укрепить нормативно-правовые основы для государственного регулирования в области аквакультуры;
- активизировать капиталоемкие научные исследования в области аква- и марикультуры, модернизировать производственные фонды (прибрежную технологическую базу, суда и орудия лова); усилить приоритетное значение аквакультуры в рамках самостоятельного национального проекта или подпроекта в национальном проекте «Развитие агропромышленного комплекса» для оказания всех форм государственной поддержки;
- обновить и реконструировать существующие производственные фонды предприятий аквакультуры.
Для обеспечения душевого потребления рыбы в соответствии с потребительской корзиной в 2015 г. необходимо произвести пищевой рыбной продукции в объеме 2,15 млн. тонн, а при рациональной норме - 2,58 млн. тонн. В настоящее время вылов рыбы в РФ стабилизировался на уровне 3,2 млн. тонн, из которых для пищевых целей используется 1,3-1,5 млн. тонн. Следовательно, производство рыбы в аквакультуре должно составить не менее 1,1 млн. тонн. Достижение указанных объемов возможно только при существенном расширении, модернизации и реконструкции материально-технической базы, что подразумевает придание аквакультуре статуса национального приоритета с обеспечением государственной поддержки и финансирования.
По имеющимся оценкам, для реализации проекта развития аквакультуры РФ на период 2007-2011 гг. требуется финансирование в объеме 91,2 млрд. рублей (примерно 3,50 млрд. долл. США), в том числе 43,2 млрд. рублей (1,65 млрд. долл. США) госбюджетное финансирование и местные бюджеты и 48,0 млрд. рублей (1,85 млрд. долл. США) - внебюджетные средства.
Ориентиры развития аквакультуры в РФ к 2020 г. включают: производство товарной рыбы в аквакультуре - 1,4 млн. тонн и марикультуре-400 тыс. тонн. Выпуск личинок и молоди ценных видов рыб в естественные водоемы к 2020 г. достигнет 80-100 млрд. экземпляров.
Экономический эффект от реализации на российском рынке продукции аквакультуры составит 25 млн. рублей (около 1 млн. долл. США) на 1 тыс. тонн. Аквакультура позволит создать в РФ до 100 тыс. рабочих мест и бупет способствовать расширению занятости в российских региона.
По данным Каспнирха за 2005 г. в Каспийском море выполнены сезонные (весна, лето, осень) тралово-акустическис и сетные съемки по изучению распределения, качественного состава и численности осетровых. Весенняя и осенняя съемки проводились на акватории западной части Северного Каспия. За период исследований проведено 581 траление 24,7- и 9-метровыми тралами и 139 постановок сетных порядков с размером ячеи от 28 до 180 мм. отловлено и проанализировано 1602 экз. осетровых: русского и персидского осетров - 1400 экз.. севрюги — 168 экз.. белуги - 32 экз.. шипа - 2 экз. В весенний период за 37 тралений и 29 постановок сетей было выловлено 4 экз. белуги. В траловых уловах ее отмечали на свале глубин Северного и Среднего Каспия. Диапазон глубин в местах лова изменялся от 4,0 до 26,0 м, температура воды - от 6,1 до 14,2 °С. Улов состоял из особей непромысловой длины. Средние размерно-весовые характеристики белуги в весенний период - 149,5 см и 16,5 кг соответственно. Высокие коэффициенты упитанности (у самцов и самок - 0,66) и зрелости (у самцов - 3.14 %; самок - 6.25 %) свидетельствовали о благоприятных условиях нагула. По результатам летней учетной съемки за 383 траления выловлено 10 экз. белуги, сетными порядками (17 постановок) - 8 экз. Единичные траловые уловы белуги отмечены почти во всех частях Каспийского моря.
Качественная структура белуги в различных частях моря (северная, средняя, южная) не отличалась большим разнообразием. В северной части Каспия (приглубая зона) доля молоди в улове составила 100 %. Длина рыб варьировала от 145,0 до 182,0 см, масса - 21,0-42,0 кг. средние показатели - 181,5 см и 4 ) ,5 кг. В средней части моря длина составила 171,0-295,0 см, масса - 42,0-175,0 кг, средние значения -232,0 см и 90,25 кг, доля молоди в этом районе не превышала 50,0 %. В южной части моря ее доля возросла до 68,4 %, а средние показатели длины и массы снизились до 144,7 см и 30,0 кг. Таким образом, прослеживается закономерность в распределении молоди и взрослых рыб по акватории моря: молодые особи нагуливаются в основном в северной и южной частях моря, а старшевозрастные группы рыб осваивают для нагула Средний Каспий. Средняя длина белуги на всей акватории Каспийского моря - 193,25 см, масса- 55,5 кг: особи промысловых размеров имели длину 214,4 см, массу - 83,1 кг, неполовозрелые особи - 158,0 см и 28,0 кг. Высокий коэффициент упитанности самцов и самок белуги (0,66-0,79) отмечен во всех районах моря, что подтверждает удовлетворительные условия нагула вида.
Данные осенней тралово-сетной съемки в западной части Северного Каспия, как и весенней съемки, свидетельствуют о единичном присутствии в траловых и сетных уловах белуги: за 72 траления и 13 постановок сетей отловлено всего 6 экз. непромысловых размеров (Журавлёва, Ходоревская, Левин и др., 2005 ).
По данным Каспнирха в 2004 г. выпуск молоди осетровых 6-ю рыбоводными заводами Севкаспрыбвода не превысил 62 % от запланированной численности, что составило 33,4 млн шт., в т. ч. .1,5 млн шт. белуги, 25,1 млн шт. осетра и 6,8 млн шт. севрюги.
Численность молоди белуги составила лишь 4,5 % от общего объема выпуска. Следует отметить, что часть молоди выращивалась при плотностях посадки в 2-3 раза ниже норматива (110 тыс. шт./ га): к примеру, на Сергиевском заводе с плотностью 30 тыс. шт./га, на Лебяжьем - 45 тыс. шт./га. Снижение плотности посадки создает более благоприятные условия питания, обеспечивающие увеличение средней массы за нормативные сроки до 5,0-6,0 г. Из рис. 1, представляющего гистограммы массы и длины этой молоди, следует, что в прудах преобладали мальки массой 3,0-4,5 г (56%). Количество молоди массой от 0,4 до 2.9 г, т. е. менее установленного стандарта (3,0 г), оказалось значительно ниже среднемноголетних значений для этого вида и составило 21% общей выборки. Достаточно многочисленной оказалась группа массой 5,0-5,5 г (17,2%). Пик-распределения длины тела белуги был у мальков от 8,5 до 10,5 см. (Григорьева, Михайлова, Крупий и др., 2004).
Глава 1. Биологическая характеристика белуги в связи со средой обитания и образом жизни
1.1 Общая биология вида
Тип: Хордовые (Chordata)
Под тип: Черепные (Craniata)
Над класс: Челюсноротые (Gnathostomata)
Класс: Рыбы (Pisces)
Над отряд: Хрящевые ганоиды (Chondrostei)
Семейство: Осетровые (Acipenseridae)
Род: Белуги (Huso)
Вид: Белуга (Huso huso)
Тело массивное, толстое (huso в переводе с латинского – свинья). Рыло короткое тупое. Рот полулунной формы, большой, но, в отличие от Калуги, не переходящий на бока головы. В спинном плавнике 48–81 лучей, в анальном – 22–41. Спинных жучек – 9–17, боковых – 37–53, брюшных – 7–14. В спинном ряду первая жучка наименьшая. Жаберных тычинок 17–36. Спина и бока тела серовато-темные, брюхо белое. Белуга (рис. 1) относится к 120-хромосомной группе осетров (Витвицкая, Тихомиров, Егоров, 2002).
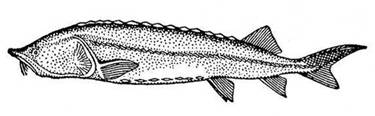
Рис. 1. Белуга
Распространение. Бассейны Черного, Азовского, Каспийского и Адриатического морей. До зарегулирования стока очень высоко поднималась по рекам. По Волге доходила почти до верховьев, встречаясь в Оке, Шексне, Каме, Суре и других притоках. По археологическим материалам вылавливалась даже в р. Москве. В настоящее время ареал ограничен нижними плотинами ГЭС. В Азовском море практически полностью исчезла, раньше, до постройки плотин, в Кубани поднималась до станицы Ладожская и, возможно, выше, в Дону – до Воронежской области. На рисунке 2 обозначены утраченные места обитания, крестиками – места археологических находок остатков белуги (Решетников, 2002).
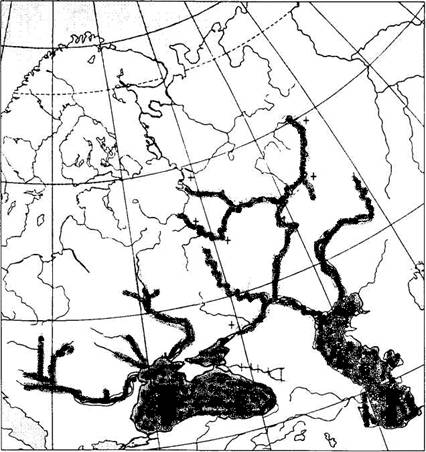
Рис. 2. Ареал обитания белуги (+ – места археологических находок остатков белуги, черная штриховка – ареал обитания белуги.)
Возраст и размеры. Одна из крупнейших проходных рыб. В прошлом достигала длины более 5 м и массы более 1000 кг. Продолжительность жизни таких крупных экземпляров, видимо, превышала 100 лет. В 1970 г. в промысловых уловах на Волге средняя длина самок белуги составляла 267 см и масса 142 кг, самцов соответственно – 221 см и 81 кг. (Решетников, 2002).
Образ жизни. Проходная рыба. В большинстве крупных рек имеет озимую и яровую формы. В Волгу входит в сентябре-октябре (озимая форма) и в марте-апреле (яровая форма). В этой реке преобладает озимая, зимующая на ямах, раса. В Урале, наоборот около 70% мигрирующих рыб составляет яровая раса, нерестящаяся в год захода в реку. Период размножения приходится на апрель-май. Нерест проходит на пике паводка и начинается при температуре воды 6–7 °С. Оптимальными температурами являются 9–17 °С. Икра откладывается на глубоких местах (от 4 до 12–15 м) с быстрым течением, на каменистых грядах и галечных россыпях. Плодовитость в зависимости от размеров самок колеблется от 200 тыс. до 8 млн. икринок. Икринки крупные, у волжской белуги их диаметр варьирует от 3,6 до 4,3 мм и масса от 26 до 36 мг. Продолжительность эмбрионального периода при температуре воды 11–12 °С составляет в среднем около 200 ч. Молодь белуги и взрослые рыбы после нереста в реке не задерживаются и скатываются в море. Половозрелость у белуги наступает очень поздно: у самцов не ранее 12–14 лет, у самок – к 16–18 годам. Нерест не ежегодный. Белуга очень рано переходит на хищное питание. В Каспийском море ее основными кормовыми объектами являются вобла, судак, сазан, лещ, кильки и другие рыбы. Белуга охотно поедает собственную молодь и других осетровых. В Черном море в ее желудках встречаются в основном хамса и бычки. Молодь белуги длиной до 5 см питается придонными беспозвоночными – мизидами, гаммаридами, олигохетами и др. (Бабушкин, 1953).
Статус вида. Ценнейшая промысловая рыба. Основным бассейном ее добычи всегда был Каспий. Еще в начале 80-х годов прошлого столетия ее здесь вылавливали от 1,6 до 2,0 тыс. т. В результате зарегулирования стока рек практически полностью прекратилось естественное воспроизводство этого вида, и ее запасы целиком поддерживались за счет искусственного разведения на рыбозаводах. В последние годы, особенно после распада СССР, численность и уловы белуги катастрофически снижаются. Белуга, как вид, находящийся под угрозой исчезновения, внесена в "Красную книгу МСОП", а азовский подвид белуги – в "Красную книгу Российской Федерации" (Решетников, 2002).
1.2 Эмбриональное развитие
Эмбриональный период развития осетровых рыб состоит из 5 этапов в 36 стадий:
1 этап – оводнение икринки и появление бластодиска (1–3 стадии).
2 этап – дробление бластодиска до бластулы (4–12 стадии).
3 этап – образование зародышевых пластов – гаструляция (13–18 стадии).
4 этап – дифференциация зародышевых пластов на зачатки основных органов (19–28 стадий).
5 этап – развитие зародышей от начала пульсации сердца до вылупления (29–36 стадии).
1. Этап (1-3 ст.)
Оводнение икринки и появление бластодиска. Выметанные в воду и тут же осемененные яйца опускаются на дно, заносятся под камни и вскоре приклеиваются студенистой оболочкой к грунту. Оплодотворение вызывает кортикальную реакцию, и между цитоплазматической мембраной яйца и оболочкой возникает узкая (в несколько мкм) прослойка выделенною материала кортикальных гранул.
Яйцо внутри высвобождается от оболочек и поворачивается соответственно положению центра тяжести, вегетативным более тяжелым полюсом вниз, а анимальным вверх. В это время в щелевидное пространство под оболочкой выделяется гидрофильный коллоид из лакун на анимальном полюсе. Коллоид, привлекая воду из окружающей среды, набухает, и возникает периветеллиновое пространство. В яйце происходит перемещение пигмента. Он стягивается к центру анимального полюса и светлое пятно исчезает. Вскоре по краю анимальной области появляется светлая полоса – светлый серп.
В это время значительно возрастает прочность яйцевых, оболочек. Увеличение прочности оболочек связано с условиями развития зародышей под камнями на течении, где икра подвергается ударом камешков, песка и других предметов. Как только кортикальная реакция осуществится и между поверхностью цитоплазмы и оболочками возникнет тонкая прослойка выделенного материала, яйцо освобождается внутри оболочек и начинает поворачиваться, в соответствии с положением своего центра тяжести, богатым желтком вегетативным полушарием вниз и анимальной областью, вверх.
Стадия 1 – оплодотворенного яйца в первые минуты после оплодотворения, когда оно по своему виду еще не отличается от неоплодотворенного яйца (рис. 3).
Пигментный рисунок анимальной области не изменен, в центре ее светлое полярное пятно; оболочки плотно прилегают к яйцу и еще не начали набухать. Через несколько минут после осеменения (от 2 до 13 мин.) наружная оболочка яиц становится клейкой и прочной.
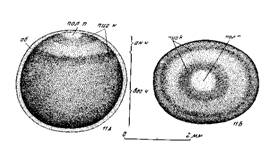
Рис. 3
Стадия 2 – оплодотворенного яйца после поворота и образования перевителлинового пространства (рис. 4).
Ко времени завершения поворота в верхней части икринки, между яйцом и оболочкой, становится различимой (если смотреть на яйцо сбоку) узкая щель, которая затем постепенно расширяется. В это время из цитоплазмы под оболочку выделяется осмотически активный коллоид (ранее содержавшийся в сети лакун анимальной области), который набухает, привлекая воду из окружающей среды; поверхность анимальной области яйца уплощается и между нею и оболочками образуется довольно значительное пространство, называемое перевителлиновым.
Своеобразный рисунок анимальной области яйца — светлое полярное пятно в центре и темные пигментные кольца вокруг него — после оплодотворения изменяется. Пигмент постепенно стягивается к центру анимальной области, образуя здесь темное скопление светлое полярное пятно, исчезает или, если оно было очень большим (что часто наблюдается у белуги), сильно уменьшается в размере.
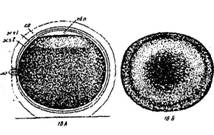
Рис. 4
Стадия 3 – светлого серпа (рис. 5). У края анимальной области нередко заметна светлая, тогда совершенно белая полоса полулунной формы (светлый серп).
Полоска эта постепенно расширяется и превращается в светлый серп. Иногда серп бывает ярким, с четкими границами, в других случаях границы его менее определенны, в некоторых партиях его совсем не удается различить.
Специально проведенные наблюдения и опыты показали, что сторона яйца со светлым серпом становится затем спинной стороной зародыша, причем воображаемая плоскость, проведенная через середину серпа и центры анимальной и вегетативной областей, соответствует будущей среднеспинной плоскости зародыша. Таким образом, зародыш приобретает двусторонне-симметричное строение в самом начале развития, еще до того как яйцо начинает дробиться.

Рис. 5
2 этап (4–12 стадии)
Дробление. Оплодотворенное яйцо многократно делится и превращается в многоклеточный пузырек — бластулу. Клетки, возникающие при дроблении яйца, называются бластомерами.
Деление яйца можно видеть даже невооруженным глазом. Для этого надо смотреть на яйцо со стороны анимальной области. Удобно наблюдать дробление на приклеенной икре.
Стадия 4 – первого деления (рис. 6). Первая борозда делит сначала анимальную область на два бластомера. Затем борозда продвигается очень медленно на вегетативную часть яйца, т. к. огромное скопление желточных зерен и жира затрудняет ее распространение.

Рис. 6
Стадия 5 – второго деления (рис. 7). Анимальная область яйца разделяется на четыре бластомера приблизительно равной величины.
Еще не успеет первая борозда достигнуть экватора, как в центре анимальной области, перпендикулярно к ней, также в виде коротких белых полосок, закладываются борозды второго деления, и постепенно анимальная область зародыша разделяется на четыре части сходной величины. При постоянной температуре борозды второго деления появляются через промежуток времени, равный одной трети периода от осеменения до появления первой борозды.

Рис. 7
Стадия 6 – Третьего деления (рис. 8 - вид сверху).
При третьем делении начинается разделение яйца на 8 бластомеров. Расположение борозд при этом бывает разлит в яйцах удлиненной формы борозды закладываются параллельно первой борозде или под небольшим углом к ней, образуя характерную фигуру в виде буквы (Ж) в яйцах округлой формы они обычно располагаются по радиусам. Рисунок, образуемый бороздами дробления, как правило, не бывает геометрически правильным, бластомеры имеют разные форму и величину.
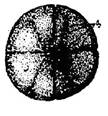
Рис. 8
Стадия 7 – четвертого деления (рис. 9). Борозды второго деления близки к смыканию; борозды третьего деления приближаются к экватору. При четвертом делении, когда в более узких бластомерах борозды располагаются горизонтально, отделяя к центру анимальной области бластомеры небольшой величины, а в более широких — косо или радиально.
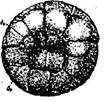
Рис. 9
Стадия 8 – пятого дробления (рис. 10 - вид сверху)
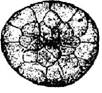
Рис. 10
Стадия 9 – седьмого деления (рис.11).
Борозды полностью разделяют богатую желтком вегетативную область. При этом четко выступает неравномерность дробления, свойственная осетровым: бластомеры, обособляющие в верхней, анимальной части зародыша, на всех стадиях дробления имеют значительно меньшие размеры, чем вегетативные бластомеры.

Рис. 11
Стадия 10 – позднего дробления (рис. 12) Последовательное деление привели к уменьшению размеров бластомеров.

Рис.12
Стадия 11 – ранней бластулы (рис. 13). В анимальной области отдельные бластомеры еще хорошо различимы, при небольшом увеличении. Внутри зародыша между бластомерами образовалась полость дробления бластоцель, зародыш приобретает строение полого шарика – бластулы.

Рис. 13
Стадия 12 – поздней бластулы (рис. 14). В анимальной области отдельные клетки при небольшом увеличении не различимы. Между мелкими анимальными и относительно крупными вегетативными бластомерами имеется зона бластомеров промежуточного размера, носящая название краевой зоны.

Рис. 14
3 этап (13–18 стадии).
Гаструляция. После того как оплодотворенное яйцо, благодаря непрерывному ряду делений, превратилось в многоклеточный пузырек — бластулу, в нем начинаются сложные перемещения: материал краевой зоны вворачивается внутрь, а светлая анимальная область разрастается и покрывает снаружи темные вегетативные клетки. Эти процессы называют гаструляцией, а зародыш в описываемый период — гаструлой.
К концу гаструляции снаружи остается только светлый материал анимальной области бластулы, который теперь покрывает всего зародыша и образует наружный слой стенки гаструлы, а краевая зона и богатые желтком темные клетки вегетативной области все оказываются внутри, где они образуют внутренний, слой зародыша.
Стадия 13 – начало гаструляции (рис. 15). Немного выше экватора на спинной стороне зародыша образовалась узкая, сильно пигментированная полоска.

Рис. 15
Стадия 14 – ранней гаструляции (рис. 16). На месте пигментной полоски образовалось углубление бластопор.

Рис. 16
Стадия 15 –средней гаструлы (рис. 17). На этой стадии светлый анимальный материал покрывает 2/3 поверхности зародыша. Бластопор замыкается в кольцо.
Рис. 17
Стадия 16 –большой желточной пробки (рис. 18). Щель бластопора замыкается в кольцо, окружая темные клетки нижней части зародыша, эта область приобретает вид темной пробки, воткнутой в светлый круглый пузырек, от чего она и получила название желточной пробки
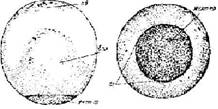
Рис 18
Стадия 17 – малой желточной пробки (рис. 19). Вся поверхность зародыша, за исключением желточной пробки, небольшого размера, покрыта светлым анимальным материалом

Рис. 19
Стадия 18 – щелевидного бластопора (рис. 20). Темный вегетативный материал полностью покрыт светлым анимальным (процесс гаструляции закончен). Края бластопора сомкнулись, между ними образовалась узкая щель

Рис. 20
4 этап (19–28 стадий).
Дифференциация зародышевых пластов на зачатки основных органов.
По окончании гаструляции у зародыша постепенно формируются зачатки основных систем органов: нервной, пищеварительной, выделительной, кровеносной, а также мускулатуры и хорды. Зародыш на этом этапе развития по-прежнему неподвижен, питается за счет желтка и дышит всей поверхностью тела. Многие из тех изменений, которые происходят с зародышем, можно проследить, наблюдая за развитием при помощи простой лупы. Покровы зародыша очень тонки и полупрозрачны, поэтому внутренние органы просвечивают через них. Обладая острым зрением, можно многое увидеть, рассматривая зародыша даже простым глазом.
Стадия 19 – ранней нейрулы (рис. 21). На спинной стороне зародыша образуется нервная пластинка. Начинают обозначатся нервные валики вокруг головного отдела нервной пластинки, они еще мало приподняты.

Рис. 21
Стадия 20 – широкой нервной пластинки (рис. 22) Нервные валики вокруг головного отдела нервной пластики четко обозначены.
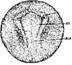
Рис. 22
Стадия 21 – начало сближения нервных валиков. Впервые обозначаются зачатки выделительной системы в виде коротких светлых тяжей, просвечивающих через покровы. Они располагаются по бокам от туловищного отдела нервной пластинки.
Стадия 22 - поздней нейрулы (рис 23). Нервные валики в туловищном отделе сближаются. Зачатки выделительной системы удлинились и видны более четко.

Рис. 23
Стадия 23 - замкнувшейся нервной трубки (рис. 24). Нервная пластинка свернулась в трубку, нервные валики сомкнулись. Шов в области их слияния имеет вид неглубокой бороздки.
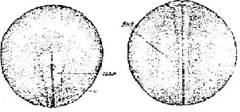
Рис. 24
Стадия 24 - появление глазных выростов и утолщения переднего конца зачатков выделительной системы (рис. 25). Шов в области слияния нервных валиков менее заметен. В задней части переднего мозгового пузыря образовались глазные выросты. По бокам от заднего мозгового пузыря возникли зачатки внутреннего уха. Впереди головного мозга, примыкая к нему, обозначается светлая пластинка — зачаток железы вылупления. В передней части закладок выделительной системы образуются утолщения.
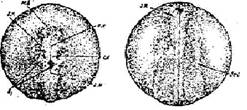
Рис. 25
Стадия 25 - сближения боковых пластинок и образования утолщения в области зачатка хвоста (рис. 26) По бокам от переднего конца головного мозга образовались зачатки обонятельных мешков. Впереди мозга менее виден зачаток железы вылупления. Боковые пластинки (из которых позднее образуется сердце, а также выстилка околосердечной полости и полости тела) достигли переднего конца головы; передние суженные выросты их сближаются. В зачатке почки дифференцируются почечные канальцы. В заднем конке зародыша возникло возвышение - еще необособленный зачаток хвоста.
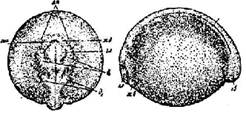
Рис. 26
Стадия 26 — слияния боковых пластинок и начала обособления хвостового отдела зародыша (рис. 27). Сблизившиеся ранее передние выросты боковых пластинок сливаются, и в месте их соединения начинается образование зачатка сердца. Петля, образуемая выводным почечным каналом, значительно удлинилась. Начинается обособление зачатка хвоста, имеющего в это время форму короткой и широкой лопасти
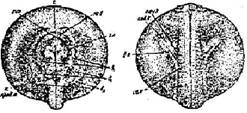
Рис. 27
Стадия 27 - короткой сердечной трубки (рис. 28). Начинается процесс обособления головы - распластанные на брюшном отделе тела зачатки головных органов начинают стягиваться к средне - спинной линии. Образовался зачаток сердца, имеющий вид короткой трубочки. Зачаток хвоста удлинился и слился.
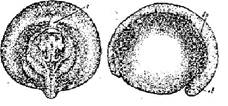
Рис. 28
Стадия 28 - прямой удлиненной сердечной трубки (рис. 29). Зародыш не подвижен, туловищные мышцы еще не реагируют на раздражение сокращением. Голова уже заметно приподнята. В связи с этим зачаток железы вылупления приобрел палочковидную форму.
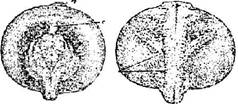
Рис. 29
5 этап (29–36 стадии).
Развитие зародышей от начала пульсации сердца до вылупления. Уловить первые сокращения сердца нелегко, так как они очень слабы и от одного сокращения до другого проходит много времени.
Но довольно скоро сокращения становятся сильнее, и сердце начинает пульсировать с определенным ритмом. Чтобы наблюдать сердцебиения, нужно положить зародыша на часовое стекло пли в чашечку и в течение нескольких минут под лупой внимательно следить за сердцем.
После того как сердце начнет пульсировать, мышцы туловища тоже приобретают способность сокращаться; если зародыша уколоть в спину иглой, замечаются слабые мышечные подергивания.
В период от начала кровообращения и до вылупления значительно изменяется внешняя форма зародыша. Голова обособляется и немного увеличивается, хвост сильно вырастает в длину, распрямляется и превращается в мощный орган движения личинки. Только брюшной отдел (желточный мешок) изменяется мало; благодаря большому количеству желтка в клетках стенки кишки он сохраняет крупные размеры.
Когда голова, раньше распластанная на желточном мешке, приподнимается, зачаток железы вылупления смещается на ее нижнюю поверхность, а сердце, на более ранних стадиях расположенное впереди головы, теперь оказывается под нею.
Продолжается развитие головного мозга и органов чувств. В том месте, где глазной пузырек соприкасается с покровным эпителием, образуется зачаток хрусталика. Обонятельные ямки углубляются. Слуховые пузырьки увеличиваются, стенка их становится тоньше; в это время они слегка просвечивают через эпителий по бокам от продолговатого мозга.
Очень заметны изменения в хвостовом отделе. Он значительно удлиняется, распрямляется и становится плоским, на спинной и брюшной сторонах его образуется широкая плавниковая оторочка. По мере удлинения хвост, будучи прижат оболочками к желточному мешку, загибается на брюшную сторону; при этом конец его постепенно достигает сердца.
Стадия 29 - образования изгиба сердечной трубки (рис. 30). Зародыш неподвижен, из – за раздражения начинает отвечать слабыми мышечными подергиваниями. Сердечная трубка значительно удлинилась и изгибается. Сердце начинает пульсировать.
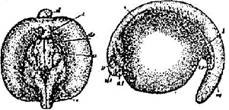
Рис. 30
Стадии 30 - конец хвоста приближается к сердцу (рис. 31). Хвостовой отдел склонен набок, хвост начал распрямляться, уплетаться, вокруг него появляется узкая, еще плохо различимая плавниковая оторочка.
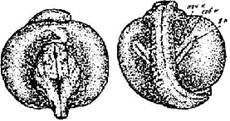
Рис. 31
Стадия 31 - конец хвоста достигает сердца (рис. 32). Хвостовой и спинной отделы наклонены набок. Если снять оболочки, зародыш делает маятникообразные плавательные движения - «плавание на месте». Хвост значительно распрямляется. Плавниковая оторочка хорошо различима. На описываемом этапе развития определять стадию удобнее всего по положению конца хвоста; нужно только иметь в виду, что между разными видами имеются некоторые отличия (так, у стерляди хвост относительно длиннее, чем у белуги, осетра и, в особенности, севрюги); поэтому следует всегда отмечать, о каком виде идет речь.
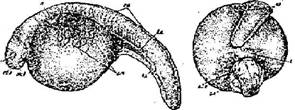
Рис. 32
Стадия 32 - конец хвоста касается головы, плавниковая оторочка шире (рис. 33).

Рис. 33
Стадия 33 - конец хвоста немного заходит за голову, достигая уровня глаз (рис. 34). Голова наклонена на бок, плавниковая оторочка хвоста расширилась. Если снять оболочки хвост полностью распрямляется.

Рис. 34
Стадия 34 – конец хвоста достигает начала продолговатого мозга (рис. 35). Зародыш активно двигается в оболочках, если оболочки снять, способен к слабому поступательному движению.
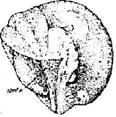
Рис. 35
Стадия 35 – начала выклева конец хвоста достигает почки (рис. 36). Выклев у белуги начинается, когда конец хвоста достигает петли, образованной собирающим и выносящим почечными каналами. В оболочках зародыш активно двигается; если их снять, плавает.
Желточный мешок имеет яйцевидную форму. В глазу может появиться пигментное пятно. Намечается ротовое углубление. Иногда обозначаются зачатки грудных плавников. Кровь бесцветная или желтовато - розовая.

Рис. 36
1.3 Постэмбриональное развитие
Период постэмбрионального развития осетровых рыб (36 – 45 стадии).
Стадия 36 - массовое вылупление (рис. 37). Форма желточного мешка яйцевидная. В глазу четкое пигментное пятно. В жаберной области обозначились складочки 2 первых жаберных карманов. На нижней поверхности головы заметно ротовое углубление. Жаберных щелей и ротового отверстия еще нет. Позади почек едва заметны зачатки грудных плавников. Кровь желтовато-розовая.
Рис. 37
Стадия 37 - появляются зачатки усиков (рис. 38); прорывается ротовое отверстие. Начинается разделение желточного мешка на желудочный и кишечный отделы. Четко выражены зачатки грудных плавников в виде небольших складочек кожи. Зачатки жаберных лепестков отсутствуют. Появляется зачаток боковой линии сейсмосенсорной системы.
Рис. 38
Стадия 38 - различимы первые меланоциты (рис. 39). Энтодермальная складка, отделяющая желудок от кишечника, неполная. Формируются первые мускульные почки в области спинного и анального плавников. Появляется зачатки жаберных лепестков на жаберной крышке и первой жаберной дуге. Боковая линия сейсмосенсорной системы достигает уровня кювьерова протока.
Рис. 39
Стадия 39 - желудок отделен от кишечника; обособились спинной и анальные плавники (рис. 40). Боковая линия сейсмосенсорной системы достигает уровня заднего грудного плавника; появляется добавочный ряд сейсмосенсорной системы.
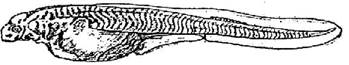
Рис. 40
Стадия 40 - различим зачаток брюшного плавника в виде узенькой складочки кожи (рис. 41). Брюшные отростки мышечных сегментов в области грудного плавника спускаются ниже его основания. Можно наблюдать первые не регулярные движения нижней челюсти. Боковая линия сейсмосенсорной системы не достигает уровня конца желудка, добавочный ряд заканчивается над грудными плавниками.
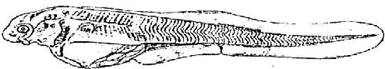
Рис. 41
Стадия 41 - края обонятельных лопастей смыкаются, но еще несращены (рис. 42). Боковая линия сейсмосенсорной системы заканчивается над спиральной кишкой, ее добавочный ряд заходит за заднюю границу грудного плавника, образуется короткий зачаток спинного ряда.
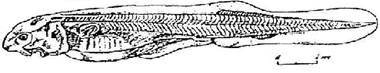
Рис. 42
Стадия 42 – появляется зачаток пилорического придатка (рис. 43).
Лопасти обонятельного органа сращены. Боковая линия сейсмосенсорной системы достигает уровня переднего края брюшного плавника: спинной ряд начинает изгибаться.
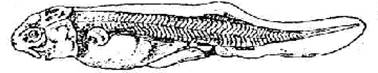
Рис. 43
Стадия 43 - рострум принимает горизонтальное положение (рис. 44).
Брюшной плавник достигает края прианальной складки. Появляются зачатки вторичных лепестков в первой жабре. Боковая линия сейсмосенсорной системы достигает уровня заднего края брюшного плавника, добавочный ряд заканчивается над спиральной кишкой, спинной ряд изогнут, и начинает расти параллельно боковой линии.
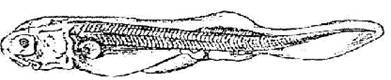
Рис. 44
Стадия 44 - основания усиков выносятся вперед, и концы их не достигают передней границы рта (рис. 45). Массовый выброс пигментных пробок. Лопасти брюшных плавников опускаются ниже края прианальной складки. В спинной плавниковой складке появляется мезенхимная полоска (общий зачаток спинных жучек). Передняя поперечная комиссура сейсмосенсорной системы переместилась дорзально и не видна с брюшной стороны; боковая линия заходит за уровень заднего края брюшного плавника, дополнительный ряд не доходит до первой границы брюшного плавника.
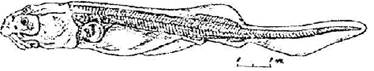
Рис. 45
Стадия 45 — перехода на активное питание (рис. 46). Видны разделительные зачатки жучек в спинной плавниковой складке. Боковая линия сейсмосенсорной системы заходит за уровень .средней части спинного плавника, спинной ряд заходит за уровень заднего края грудною плавника.

Рис. 46
Личиночный период развития осетровых рыб.
В личиночном периоде обычно выделяют два этапа:
I личиночный этап - этап смешанного питания.
Дыхание становится наружно-жаберным. Появляются зачатки брюшных плавников. Рот нижний, перед ним четыре усика. Голова большая Рыло постепенно становится заостренным. Плавниковая кайма на месте непарных плавников обособляется. Хвостовой плавник большой, гетероцеркальный. Вдоль спинного края плавниковой складки начинаются появляться бугорки - зачатки жучек. Сначала они еще не выходят за пределы складки. На верхней челюсти закладываются зубы. Питание у зародышей смешанное, этап длится 3 дня, анальное отверстие замкнутое, у личинок сформированное. Меланин, имевшийся в кишечнике зародыша выделен.
II личиночный этап - экзогенного питания.
Желточный мешок полностью резорбировался, питание экзогенное. Рот личинки при открывании выдвигается, но ротовой трубки, типичной для более позднего малькового этапа еще нет.
Увеличивается число и длина жаберных лепестков. Жаберная крышка разрастается, покрывая собой наружные жаберные лепестки почти до трети их длины. Образовался спиральный клапан. Длина личинок 21 мм.
Мальковый период развития осетровых рыб.
Личиночные плавниковые складки резорбируются. Вентральная поверхность туловища малька становится плоской, имеются ряды спинных, боковых и брюшных жучек. На голове развиваются покровные типы. Жучки и костные пластинки образуют на теле малька защитный и колючий панцирь. Жаберные крышки разрастаются полностью прикрывая жаберные лепестки. В отличие от личинок челюсти малька не имеют зубов. Зубы исчезают тогда, когда жаберные крышки начинают функционировав как всасывающий аппарат. При этом утрачивается не