


Биологическая эволюция и морфогенез
С.Б. Пашутин
Биологическое разнообразие нашей планеты недвусмысленно указывает на успехи биологической эволюции. И хотя это стадия эволюции, качественно отличная от химической, ее общие принципы остаются теми же и явственно проступают на всех ступенях филогенеза. Прежде всего это относится к физико-химическим особенностям молекулярного взаимодействия. Если в условиях ограниченности ресурсов на образование какого-либо вещества тратится меньше времени, то это вещество и становится доминирующим в конкретной ситуации. Скорость реакции является признаком, по которому она отбирается в ходе эволюции из множества других сходных химических реакций.
Сходство биогенных и абиогенных форм материи особенно заметно в способности изменять свои свойства в зависимости от условий среды. Эта способность проявляется в виде переходов между устойчивыми состояниями системы при воздействиях факторов окружающей среды, если только они не приводят к разрушению системы.
Биологические системы являются открытыми нелинейными системами, способными к самоорганизации. Они функционируют за счет поступления энергии извне. При недостатке энергии они становятся менее организованными и могут самопроизвольно разрушиться. Организация, т.е. снижение собственной энтропии системы, происходит за счет повышения энтропии окружающей среды. Разрушение системы также сопровождается повышением энтропии. Можно сказать, что разрушение происходит само собой, а любое созидание требует затраты энергии.
Именно на этих фундаментальных основах природной целесообразности и происходило последующее усложнение и совершенствование исходно примитивной биологической системы.
Почему в ходе эволюции появился необычайно широкий ассортимент различных форм жизни? В принципе, могла бы существовать одна-единственная открытая биологическая система, поскольку в каждой из ныне существующих популяций как основа, так и само качество жизни, в целом одинаковы.
Возможно, так оно и было сначала. Среди первых представителей жизни на Земле, так называемых протобионтов, скорее всего особого разнообразия не было. Разнообразие сложилось постепенно, когда в результате накопления биомассы первичных организмов стало возможным освоение все большего пространства. И уже в зависимости от разных условий местообитаний у протобионтов могли возникать зачатки внутривидовой изменчивости по морфофизиологическим показателям.
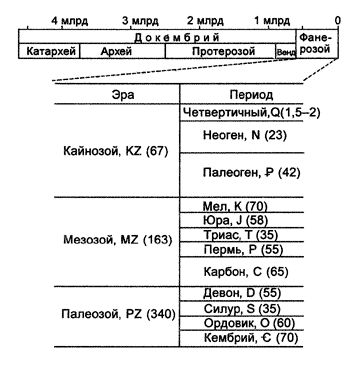
Интересно, что этот эволюционный период оказался наиболее продолжительным, заняв едва ли не 85% времени всей биологической эволюции. Так, если самые древние одноклеточные организмы появились приблизительно 3, 5 млрд лет назад, а Земля образовалась за 1–1, 5 млрд лет до возникновения первых устойчивых форм жизни, то все многообразие живой природы сформировалось в кембрийском периоде палеозойской эры, т.е. 530–540 млн лет назад и за исторически короткий по сравнению с предыдущими архейской и протерозойской эрами срок в 5–10 млн лет.
Понятно, что кембрийский «скачок» был обусловлен комплексом значимых для эволюционного развития изменений условий. Прежде всего это тектонические сдвиги и перемещения материков, изменивших ось вращения нашей планеты, а также накопление в земной атмосфере кислорода, что благоприятствовало переходу к более эффективному для жизнедеятельности аэробному метаболизму. Кроме того, увеличение концентрации кислорода способствовало снижению интенсивности ультрафиолетового излучения, что позволило протобионтам заселять не только глубины океана, но и осваивать иные ареалы.
Протобионты оказались в непривычных для них условиях внешней среды, как географических, так и физико-химических, зависящих от геологических характеристик конкретных районов. Подобные стрессорные воздействия, произошедшие незадолго до начала кембрийской эпохи, явились для древних форм жизни мощным стимулом и дали толчок появлению новых форм.
Следует иметь в виду, что это было возможно лишь при наличии накопленных точечных или хромосомных мутаций, зафиксированных в геноме. Расширению генома могла способствовать вставка в него чужих генетических текстов, например при проникновении в клетку ретровирусов, а также реализация разнообразных механизмов образования полиплоидных форм. Генетическое разнообразие эукариот, имеющих оформленное ядро, могло достигаться с помощью хромосомных рекомбинаций, поскольку ранние признаки полового диморфизма начали появляться (в частности, у водорослей) еще задолго до кембрия, более 2 млрд лет назад.
Если бы условия существования оставались неизменными, то накопленные генетические изменения никогда бы не реализовались, поскольку биологическая структура для существования в тех или иных условиях задействует вполне определенный и оптимальный набор свойств, и поэтому ни в каких генетических вариациях не нуждается.
Пример с aэpoбным метаболизмом в условиях, когда появился кислород, убедительно демонстрирует ограниченность, неэффективность анаэробного обмена веществ, который доминировал у господствовавших в докембрийскую эпоху форм бактерий и водорослей.
Анаэробиоз тормозил эволюционное развитие (во всяком случае, не способствовал ему). Но это не значит, что в течение этой длительной филогенетической стадии в геноме одноклеточных не сумели накопиться потенциально правильные генетические сочетания, которые затем стали нужными в новых обстоятельствах, в частности в условиях аэробного окисления.
Это не противоречит генетическим законам, поскольку мутации, в том числе нейтральные, наследуются. Многие из них ничего не меняют в структурах белков, т.е. ни к каким последствиям для организма в данных конкретных условиях они не приводят. Кроме того, мутация может быть скрыта, оставаясь в пределах видовой нормы, при точечных заменах нуклеотидов. Например, изменение кодирующего аминокислоту кодона может и не повлиять на свойства белка, если оно не затрагивает активного центра фермента, состоящего, как правило, из небольшого числа аминокислот.
В то же время вполне возможно и специфическое для определенного клеточного генома проявление скрытых до поры мутаций при стрессе. Это уже приведет к ускоренному формированию новых признаков, обусловленных не одновременным возникновением новых мутаций, а ранее накопленными изменениями в генетическом аппарате.
Здесь прослеживается явная аналогия с химической эволюцией, когда возникновению жизни предшествовал длительный период накопления потенциально возможных сочетаний различных химических соединений друг с другом, необходимых для всех структурных элементов новой системы. Ход эволюционного развития нашей планеты определялся сменой условий на ней, под которые подстраивались все физические и химические процессы. Можно представить эволюционный процесс как предварительное и длительное накопление потенциально полезных химических реакций и соединений веществ с образованием структурированных систем, которые ожидают подходящих условий, чтобы оказаться востребованными.
Эволюция часто прибегает к перебору вариантов, хорошо себя зарекомендовавших во многих системах. Об этом наглядно свидетельствует сходство молекулярных механизмов, а в ряде случаев и функций, у разных организмов. Некоторые из них в практически неизменном виде сохранились на всех иерархических уровнях эволюции – от самых древних синезеленых и пурпурных бактерий до современных млекопитающих. Так, например, в системах переноса электронов и окислительного фосфорилирования задействованы по сути аналогичные и лишь незначительно модифицированные белки-переносчики: светочувствительный белок сетчатки глаза (родопсин), фотосинтетические хромофоры архебактерий (бактериородопсин), пигменты водорослей (фукоксантин диатомовых и бурых или фикоэритрин красных водорослей) и растений (хлорофилл), гемоглобин эритроцитов или мембранные митохондриальные белки, участвующие в процессах клеточного дыхания (цитохромы) и т.д.
Таким образом, докембрийский этап филогенеза можно рассматривать как чрезвычайно важную стадию биологической эволюции, несмотря на то, что в течение достаточно продолжительного времени никакие другие обитатели, кроме одноклеточных бактерий и водорослей, на Земле не существовали. Его историческая ценность заключалась в накоплении «биосырья» в виде самых разных молекулярных сочетаний для последующего построения уже консервативных структур, значимость которых закреплялась последующим отбором. Кроме того, эта длительная стадия эволюции послужила полигоном для отлаживания прогрессивных приспособительных биологических технологий.
Эволюционное усложнение организмов связано с совершенствованием репродуктивных и морфогенетических механизмов, приведшим к принципиально новому этапу развития, в результате которого и возникло наблюдаемое видовое разнообразие. В плане оптимизации адаптационных возможностей биосистемы процессы размножения и формообразования достаточно тесно связаны друг с другом, несмотря на различные мотивы их возникновения.
Вероятно, феномен клеточного деления (более уместно говорить об умножении, или копировании, клеток) не был обусловлен способностью к матричному самовоспроизведению, присущему живым системам. Скорее всего, необходимость в копировании клеток возникла в связи с несовпадением скорости роста поверхности и объема – увеличение объема клетки было лимитировано ее мембраной.
Очевидно, что скорость накопления биомассы свидетельствует об эффективности поглощения исходных ресурсов и, соответственно, о большей, по сравнению с конкурентами, интенсивности заполнения жизненного пространства. Естественно, что в подобной ситуации самоорганизующаяся система совершила закономерный переход к клеточному дроблению. Он же был удачно использован и для переноса наследственного материала от материнской особи дочерним клеткам, поскольку никакие другие способы для этого не подходят, за исключением более затратного, с точки зрения биологической целесообразности, вирусного механизма передачи генома.
Но множество отдельных разобщенных клеток, даже адекватно приспособленных к существованию в конкретных условиях местообитания, – это еще не многоклеточная структура. Отсутствие значимых для эволюционного усложнения резких изменений внешних условий в течение большей части докембрийской эпохи не оставляло одноклеточным организмам практически никаких шансов на выраженную клеточную специализацию, обрекая их на физиологическую адаптацию, необходимую для конкуренции за ресурсы в своей узкой экологической нише. Тем не менее, ближе к концу протерозойской эры накопление определенного генетического фонда в сочетании с некоторыми изменениями условий внешней среды облегчило переход к различным новым формам жизни.
Постепенно стали появляться разные типы автотрофов, самостоятельно синтезирующих все необходимые питательные вещества, виды сапрофитных гетеротрофов, разлагающих органические остатки погибших организмов, что, несомненно, способствовало эволюционному оживлению. Особенно отличились представители простейших, поскольку до появления первых хищников – инфузорий, амеб и жгутиковых – на популяции древних одноклеточных никто не посягал.
По-видимому, новые обстоятельства благоприятствовали формированию многоклеточных структур как более мобильных и лучше приспособленных к выживанию в изменившихся условиях существования. Им было легче противостоять внешней агрессии, проще добывать ресурсы, т.е., в целом, они оказались гораздо устойчивее своих одноклеточных собратьев.
Наряду с экологическими причинами развитие многоклеточности могло быть опосредовано проявлением генетических аномалий. Так, например, нельзя исключить, что конгломерат из не полностью разошедшихся при митозе клеток мог быть стадией, предшествующей более организованной биологической форме в виде упорядоченной многоклеточности.
Структурное усложнение допускало клеточную специализацию развития, недоступную одноклеточным организмам. Хотя парамеции, миксобактерии, слизевики миксомицеты и достигли вершин дифференцировки, но в достаточно узком диапазоне и на уровне примитивных реакций в силу ограниченных возможностей автономной клетки.
Создание сверхчувствительных органов с низким порогом восприятия разнообразных стимулов окружающего мира является прерогативой исключительно высокоорганизованных структур. Более совершенные многоклеточные организмы пришли к этому путем совершенствования функциональных структур своих одноклеточных предшественников. Так, первые зачатки специализации можно наблюдать у вольвокса – колониальной формы жгутиковых. В клетках переднего сегмента этой шарообразной структуры расположены крупные светочувствительные стигмы, тогда как на обратном полюсе находятся клетки, предназначенные для размножения.
Фактически, природа воспользовалась оптимальным алгоритмом организационного развития, основанном на фрактальном принципе внутреннего подобия. В определенной степени на это указывает биогенетический закон Геккеля–Мюллера о повторяемости стадий предков при индивидуальном зародышевом развитии современного организма. Прохождение через все уровни эволюционной иерархии сходных принципов структурного построения свидетельствует о заложенных в основе биологического развития фрактальных механизмах, видимо, как самых лучших или же наиболее подходящих вариантах морфогенеза. Действительно, изначальная жизненная форма в виде сферической полости в том или ином виде воспроизводится на всех этапах филогенеза. Устойчивая полостная конструкция, оказавшись удачным решением на уровне одиночной клетки, сохраняется и у многоклеточных организмов. Стремление к обособлению от окружающей среды становится более совершенным, а внешняя оболочка, защищающая внутреннюю полость, выстраивается организмом уже из собственных специализированных клеток.
К эукариотической клетке, тем самым, применим двойной стандарт: ее можно рассматривать и как самостоятельное целостное образование, и как структурную единицу высокоорганизованной биологической системы. Подобное делегирование полномочий широко задействовано в морфогенезе, что позволяет еще не дифференцированным стволовым зародышевым клеткам трансформироваться в многочисленные клетки разных типов.
В эволюционном плане – это шаг вперед, но при этом каждая из специализированных клеток многоклеточного организма теряет свою, пусть и примитивную, универсальность. Такие издержки специализации не позволяют, например, многоклеточному организму размножаться простым делением.
Дифференцировка стволовых клеток в ходе онтогенеза сопровождается инактивацией или потерей определенных генных локусов. При далеко зашедшей специализации у одних клеток в той или иной степени может наблюдаться деградация ДНК, у других – полное репрессирование генома, вплоть до разрушения клеточного ядра. Поэтому, на каждой стадии специализации стволовая клетка теряет свою тотипотентность, т.е. способность реализовать всю генетическую информацию, заложенную в ядре, а ее дифференцировка становится необратимой.
Правда, существуют и исключения: растительная клетка не расстается со способностью к универсальной трансформации. У животных стромальные клетки костного мозга, не являясь тотипотентными, способны заменять погибшие специализированные клетки в разных органах. Кроме того, в экспериментах по клонированию были подобраны условия, при которых цитоплазма ооцита могла перепрограммировать ядро соматической клетки, возвращая ей тотипотентность, т.е. клетка вновь обретала универсальность стволовой клетки.
Параллельно с морфологической специализацией шло усложнение и механизма размножения, основной смысл которого – обеспечить открытой биологической системе устойчивость и предсказуемость ее развития на протяжении как можно более длительного срока. Как правило, открытая система является неравновесной и нелинейной. В некоторых ситуациях она бывает лишена устойчивости линейных систем, и тогда исходно незаметные различия в начальных условиях способны привести к совершенно неожиданным результатам.