


Базовый интегральный модуль неокортекса. Проблема и решение - дополнительный подход
С.А. Чебкасов и Л.А. Кондратьева, В.М. Корзак
НИИ нейрокибернетики РГУ им. А.Б. Когана, г. Ростов-на-Дону, Россия
В слое три неокортекса грызунов на тангенциальных срезах плаща коры мозга цитохромоксидазные "пэтчиз" размером около 200 мкм обнаруживаются всюду, включая проекционные зоны: зрительную ("блобс"), соматосенсорную (в том числе - "бочонки"), слуховую кору, а также моторную кору, и премоторную кору, ассоциативные поля 5 и 7.
Таким образом, на примере коры мозга грызунов выявлены универсальные базовые интегративные модули строения неокортекса.
Обнаруженный базовый интегративный модуль не соответствует представлениям о "колонках", как универсальном модуле, ни в одной из известных нам интерпретаций: Маунткастла, Эдельмена, Хьюбела и Визла, Гроссберга, Кройтцфельдта, Альбуса, Сентаготаи, Экклса - он имеет принципиально иную, чем традиционно предполагали, конструкцию и не может быть адекватно описан в терминах иерархии рецептивных полей или термодинамики модулей, "нейродарвинизма", но требует квантово-волнового описания.
Результаты наших физиологических и морфологических исследований и вытекающая из них наша концепция информационно-структурного дополнения дают новый взгляд на проблему базового модуля неокортекса, который находится в согласии со строго установленными фактами целого ряда экспериментальных и теоретических работ, модельными исследованиями; законами фило-, онто- и эмбриогенеза, принципами сравнительной нейроморфологии.
Полученные факты вместе с результатами прежних наших исследований и теоретических обобщений в области электрофизиологии и морфологии позволяют сделать вывод, что базовый интегративный модуль неокортекса млекопитающих найден.
С пионерских работ Лоренте де Но, который представлял себе неокортекс как совокупность дискретных, но взаимосвязанных единиц - колонок, вопрос о структурно-функциональной единице неокортекса находился в поле зрения нейрологов.
В 1957г. В. Маунткастл привел физиологические доводы в пользу существования физиологических проекционных колонок в соматосенсорной коре, составленных из функционально сходных нейронов. Их наличие он связывал с локальными терминалями специфических афферентов и с латеральным торможением. Такие проекционные колонки служили базой топической организации коры.
В 1962г. Шейблы, исследуя особенности ветвления неспецифических афферентов, ввели представление о неспецифическом модуле коры и предложили гипотезу, по которой перекрытие локальных специфических афферентов и неспецифических афферентов создают базу для колонок Маунткастла.
В 1962г Хьюбел и Визел выявили в зрительной коре мозга колонки по ориентации (полосы) - клетки колонки "предпочитали" одинаковую ориентацию полосы, но это были колонки, в которых осуществлялась последовательная интеграция входных сигналов на различных по уровню конвергенции нейронах. В отличие от колонок Маунткастла ориентационные колонки Хьюбела-Визла не были проекционными - с одного и того же локуса сетчатки в зависимости от ориентации "запускалась" та или иная ориентационная колонка., набор которых обслуживал все ориентации - гиперколонку, Эти же авторы обнаружили глазодоминантные колонки, для которых нашли морфологическую базу - чередование зон ветвления специфических афферентов, несущих сигналы от левого и правого глаза. Пространственно эти колонки перекрывались, вместе составляя повторяющийся в коре паттерн, который соответствовал единице топической организации - проекционной колонке. Такие блоки коры Хьюбел и Визл обозначили как модуль и предложили его модель - известный (и критикуемый) "кристалл льда".
Попытки обосновать морфологически существование колонок по ориентации полос на базе исследования активности деоксиглюкозы нельзя считать удовлетворительными с точки зрения выявления дискретных блоков коры мозга, что сами авторы позже фактически признали.
Между тем, число колонок, выявляемых в зрительной коре все нарастало: колонки по глубине восприятия, цветовые колонки, колонки по дирекциональности, колонки по оси движения, колонки по пространственной частоте и т.д.
Однако в центре внимания были именно колонки по ориентации (полосы) из-за того, что с ними Хьюбел и Визл, а вслед - многие, связывали базовую функцию зрительной коры. Парадоксальным образом в зрительной коре грызунов долгое время не удавалось выявить колонок по ориентации (а это 2/3 всех млекопитающих), но удалось обнаружить свидетельства колонок на освещение диффузным светом, вспышкой. Наиболее доказательно это было сделано в 1972-1973гг. в работах Л.Н. Подладчиковой. Причем, для таких колонок обнаруживались и морфологические корреляты в связи с выявлением (М. Тюрбеева, 1973), имеющих ту же размерность, что и колонки на диффузный свет на плане коры повторяющихся групп пирамидных нейронов слоя V.
Со временем колонки на диффузный свет были выявлены и в коре приматов. Ими оказались"blobs", обнаруженные в 1978г. по активности цитохромоксидазы. Хьюбел, который ввел для колонок зрительной коры с повышенной активностью цитохромоксидазы обозначение "blobs" ошибочно считал их принадлежностью зрительной коры лишь приматов. Важный шаг в изучении таких колонок, получивший затем развитие в ряде работ, был сделан этим знаменитым ученым с коллегами, когда им удалось показать адресную организации ассоциативных связей применительно к "blobs" и колонкам по ориентации (полос).
В 1970г. в соматосенсорной коре Велкером и в 1971 Вулси были выявлены "бочонки", необычайно яркие морфологически структуры, которые принято рассматривать в качестве классических проекционных колонок, но оказавшихся, однако, как это показал, видимо впервые, А.Г. Сухов с сотрудниками: В.Дуканичем, Е.Н. Стадниковым и В.А.Н. Ласковым все же не проекционными колонками Маунткасла (А.Г. Сухов придерживается противоположной точки зрения), но модулями в смысле Хьюбела, Визла.
Большую роль в выявлении модульной организации небочонковых зон соматосенсорной коры, и, что особенно важно - ассоциативных зон неокортекса грызунов сыграли работы А.С. Батуева и Бабминдры, а для высших млекопитающих - Казакова и Кузнецова. Все эти работы были обобщены со временем в монографии А.С. Батуева, который ввел понятие структурного модуля, и создал представление об их иерархии.
В целом в результате интенсивных исследований колонок на протяжении более чем полувека накопился весьма разношерстный, во многом противоречивый материал, который нуждался в обобщении.
Атака была предпринята с двух сторон - морфологом и физиологом.
В. Маунткасл - "отец" физиологических колонок - построил модульную концепцию мозга, в базу которой положил представление о миниколонке - универсальной единице неокортекса с единым клеточным составом, которая содержит и основной набор локальных цепей переработки информации. Эту идею на базе эмбриологических исследований поддержал (справедливо критикуемый) Ракич, а Эдельмен на этой базе построил концепцию нейродарвинизма.
Несмотря на жесточайшую критику (особо отметим статью "нейроэдельманизм) Маунткасл и Эдельмен, кажется, по-прежнему придерживаются своих взглядов.
Не менее поразительно и то, что при интенсивных исследованиях нам не известны (одно исключение) экспериментальные работы, где бы пространственное разрешение применяемых методов соответствовало одной миниколонке.
Между тем идея универсальной базовой колонки противоречила данным морфологии о тангенциальной гетерогенности неокортекса, в том числе, - особенно доказательным работам Антоновой, показавшей, что в коре соседние миниколонки весьма различны по базовому клеточному составу: миниколонки, где преобладают пирамидные клетки, чередуются с теми, где преобладают звездчатые нейроны. Такие миниколонки (как это несомненно для нейролога) должны различаться и функционально. Совершенно наглядно все это на примере "бочонков", в том числе, - из работ Т.А. Лапенко и А.Г. Сухова.
Атака со стороны морфологии была предпринята Я. Сентантаготаи - сначала концептуально (концепция универсального морфологического модуля неокортекса), затем - путем обобщения исследований по адресной организации внутрикорковых связей.
В результате микроучастки коры мозга размером около 200мкм с адресными внутрикорковыми входами, выявляемые в мозге от мыши до человека, с пионерских работ Сентаготаи,1978, рассматриваются как универсальные модули неокортекса, в том числе - в работе знаменитого Экклса,1984.
В зрительной коре по мысли Сентаготаи и других авторов вслед такой модуль отождествлялся с ориентационной колонкой Хьюбела и Визла. Неслучайно поэтому Сентаготаи отрицал топографическую связь терминалей специфических волокон с отдельными модулями. Морфологически это представить вообще нелегко с учетом того, что специфическое волокно перед входом в кору разветвляется, оканчиваясь в разных топографически зонах, а размер терминалей в среднем совершенно несопоставим с поперечным размером отдельного модуля - в 200мкм. В настоящее время более подробный анализ показал, что локальные специфические терминали с поперечником как раз в модуль обеспечивает система так называемых X-клеток). Попытка представить модульную организацию неокортекса привела Сентаготаи и Эрди к концепции модульной термодинамики. А также, как мы упоминали выше, - послужила базой для теоретических представлений Эклса.
Все же охватить наличный материал непротиворечиво не удалось.
Во-первых, морфологически выявляется не один. Но минимум, два типа модулей, разнящихся по числу входящих в них клеток на несколько порядков (этот вопрос рассматривался, в частности, в работе Чебкасова).
Во-вторых, согласно работам нашей отечественной школы сравнительной морфологи, в частности - Богословской и Полякова, нервные структуры развиваются путем надстройки - так, что прежние структуры не элиминируются. Отсюда следует, что одни и те же модули должны обнаруживаться и у грызунов, и у высших млекопитающих. К той же мысли подводят выводы работы Диамонда и Хола, которые показали, что различные отряды млекопитающих произошли от общего предка и потому имеют общий план строения коры мозга с единым базовым модулем. Для зрительной коры мозга, казалось бы, это должна быть колонка по ориентации (полос). Но как уже отмечалось, долгое время данные о таких колонках у грызунов отсутствовали.
В-третьих, критерию адресных ассоциативных связей соответствует не одна, но две системы "элементарных" колонок: одна из которых - колонки на диффузный свет (эти колонки обнаружены экспериментально у всех млекопитающих), другая - колонки по ориентации (полос) - основного претендента на роль универсального модуля в зрительной коре.
С 1969г. мы исследовали конструкцию колонок на диффузный свет на примере зрительной коры мозга морской свинки, в то же время анализируя результаты исследований (по сути они оказались аналогичны) на зрительной коре белых крыс, также проводимых в НИИ НК РГУ с целью решения проблемы элементарного вероятностно-статичтического ансамбля (А.Б. Коган, О.Г. Чораян). При этом были разработаны методы с пространственным разрешением в одну миниколонку.
В 1971г. мы открыли факт полярной векторной "он-офф" организации колонок на диффузный свет.
Затем в 1978г. совместными с Л.Н. Подладчиковой и Г.Г. Бондарь был открыт факт адресной организации гетерогенных входов специфической, ассоциативной и неспецифических систем афферентов применительно к территориям отдельных колонок на диффузный свет и дана (Чебкасов) его морфологическая интерпретация, позволившая уже обосновано говорить о таких колонках как не только функциональных, но и структурных единицах зрительной коры. Подтверждение в работах А.Г. Сухова с Т.А. Лапенко факта адресной конвергенции различных афферентаций применительно к бочонковым колонкам позволило нам укрепиться в предположении (Чебкасов,1972), что мы имеем дело с универсальным структурным модулем неокортекса.
В работах Чебкасова1990-2003гг. было показано, что колонки по ориентации полос (аналоги колонок Хьюбела-Визла у высших млекопитающих) - это выходные зоны композитных полярных модулей, составленных из пары базовых "он-офф" модулей с противоположной ориентацией осей "он-офф"-диполей.
В совместных опытах с Л.Н. Подладчиковой и Г.Г. Бондарь при анализе топографии фокальных вызванных потенциалов, вызываемых стимуляцией вспышкой диффузного света, были получены первые свидетельства того, что колонки на диффузный свет с поперечным размером около 200мкм образуют объединения старшего порядка. Дальнейшие исследования с применением локального освещения и ориентированных полос позволили подтвердить сложившуюся у нас гипотезу "ворот" о роли полярной векторной "он-офф"организации колонок на диффузное освещение как конструктивного фактора, определяющего ориентационную чувствительность таких колонок и установить, что выявляемые по фокальным потенциалам макроколонки являются аналогом гиперколонок - модулей Хьюбела-Визла. В этом нашем заключении существенную роль сыграла работа Л.Н. Подладчиковой и Беляковой, показавших, что на отдельных колонках на диффузный свет конвергируют входы и от левого, и от правого глаза (база для глазодоминантых колонок). С учетом данных, полученных в нашем институте для белой крысы это позволило показать, что план строения зрительной коры мозга грызунов и приматов един.
Подробно исследовав конструкцию колонок на диффузный свет, обладающих ориентационной чувствительностью, мы с одной стороны получили решающие доказательства их полифункциональности (это было сформулировано нами уже в 1972г., а морфологами - в уже цитированной работе Богословской и Полякова - как общий принцип полифункциональности интегративных структур мозга). С другой стороны были получены данные, позволяющие рассматривать изучаемые структурно-функциональные единицы коры в качестве элементарных интегративных модулей.
При этом стало понятно, что из-за наличия как минимум двух систем модулей, необходимо уточнить термин "универсальный модуль" неокортекса до определения "базовый универсальный интегративный модуль" неокртекса.
К 1987г. мы сформулировали представление о том, что ориентационно-чувствительные колонки грызунов есть аналоги не колонок по ориентации, но "blobs".
В 1987г. мы изготовили препараты по методу Е.Н. Стадникова. Такие препараты из-за отсутствия извилин у грызунов позволяли получать тангенциальные срезы на уровне слоя Ш, включающие практически весь плащ неокортекса. Используя бинокулярный микроскоп на малых увеличениях, уже на неокрашенных препаратах, и на препаратах окрашенных на крезил-виолет и гематоксилин-эозин, мы визуально обнаружили то, что искали - группировки размером около 200мкм (за исключением аналогичных структур, но более крупных, и упорядоченных в ряды - в бочонковом поле соматосенсорной коры). Описываемые структуры выявлялись не только в проекционных зонах, включая слуховую кору, и моторной коре, но и в ассоциативных зонах (поля 5и7), а также в премоторной коре. Это давало сильный довод в пользу того, что изучаемые нами колонки в зрительной коре и те, изучаемые в нашем институте А.Г. Суховым с сотрудниками - бочонковые колонки суть частные случаи базовых универсальных модулей неокортекса. Последнее заключение усиливалось установленнием А.Г. Суховым с В.М. Дуканичем, Е.Н. Стадниковым и В.Н. Ласковым факта, что в слое IV нейроны противоположных стенок "бочонка" характеризуются противоположной дирекциональной чувствительностью. Это нам позволило сделать заключение, что и "бочонки" представляют собой полярные векторные структуры, а вместе с аналогичными данными литературы - о том, что полярная векторная организация является принципом организации базовых универсальных интегративных модулей неокортекса.
Для ориентационно-чувствительных колонок по диффузному свету нами в 1980г. было сформулировано представление, что в силу их конструкции процесс нервной интеграции не может протекать иначе, чем в виде апериодических эпизодов миллисекундной синхронизации спайков. Каждый такой эпизод - суть элементарный вероятностно-статистический нейронный ансамбль - ансамбль А.Б. Когана. Каждый такой ансамбль объединяет активность части (многие десятки и сотни) гетерогенных и полярных элементов. Это, однако, был не эргодический ансамбль О.Г. Чораяна и Сентаготаи-Эрди, но квантово-волновой. В последовательности таких ансамблей на языке временного кода содержится сообщение о результатах достигнутого в колонке уровня интеграции гетерогенных и полярных ее элементов - для иных нервных структур и к эффекторам. Сведения литературы и данные, полученные в нашем институте при изучении "бочонковых" колонок, позволяли обобщить представления о роли миллисекундной синхронизации до общего принципа функционирования интегративных структур неокортекса.
К 1990г. мы экспериментально показали, что ориентационная чувствительность колонок на диффузный свет - это не чувствительность к ориентации полосы, но чувствительность к ориентации контрастной кромки, реализуемой (в соответствии с прежними нашими представлениями) на основе структурно предопределенного асимметричного растормаживания (но не асимметричного торможения) и на основе миллисекундной синхронизации.
Тем самым, с учетом данных литературы, мы продемонстрировали обоснованность представления о том, что не колонка по ориентации полосы, но структуры, аналогичные "blobs", служат базовым элементом для миллисекундной синхронизации спайков.
Также было подтверждено предположение о композитных полярных векторных модулях (составленных из пар базовых модулей с противоположной ориентацией оси "он-офф"-диполей и соответственно "предпочитающих" противоположную ориентацию контрастных кромок) как модулях для выявления ориентации полос.
На базе полученных данных мы развили принципиально новую концепцию нервной интеграции (Чебкасов,1989-2003).
Отчетливо понимая, что ее экспериментальная база неполна, и что 2/3 млекопитающих - это грызуны, мы в 2001г. приступили к прямому выявлению "blobs" у грызунов. Для этого мы использовали те самые методы, которыми "blobs" были выявлены сначала в мозге приматов, затем - хищников, а далее - цитохромоксидазных "пэтчиз" в соматосенсорной коре грызунов, соответствующих "бочонкам, а также в слуховой коре.
Результаты этой работы перед Вами.
Методика
В отличие от "блобс" приматов, разделенных протяженными промежуточными зонами мы заранее знали (по результатам предыдущих исследований, что эти зоны значительно уже (Чебкасов,1998). Соответственно необходимо была особая точность резания при изготовлении тангенциальных препаратов слоя Ш. Здесь нам на помощь вновь пришел метод резания, разработанный Е.Н. Стадниковым. Если бы не этот метод можно было заранее ожидать тех же результатов, что получили для грызунов Хьюбел с сотрудниками, которые не смогли выявить "блобс" у грызунов. - Причина в том, что узкие промежуточные зоны между "блобс" будут выявляться на плане коры лишь тогда, когда будут ориентированы по нормали к плоскости среза. При скосе, с учетом толщины среза в 75мкм, мы неизбежно получили бы равномерное распределение окраски - промежутки между "блобс" в проекции коры оказались бы закрыты для наблюдения.
Опыты были проведены на коре мозга шести белых крыс весом около 300Г. Об активности цитохромоксидазы судили путем анализа тангенциальных срезов плаща коры мозга, приготовленных по методу Е.Н. Стадникова (Cтадников, 1986) и окрашиваемых по классической методике (Leventhal et al,1995,Marphy et al,1995), с помощью которой были выявлены цитохромоксидазные "блобс" и "пэтчис" ранее Использовались препараты фирмы ICN (USA). Фотографии срезов делались с помощью фотонасадки через микроскоп МБС-2 при увеличении 2х8.
Результаты и заключение
На рис.1 представлена фотография тангенциального среза, где видны "блобс" зрительной зоны коры мозга. А - малое увеличение. Б - большое увеличение.


А Б
На базе эмбриологических исследований зрительной коры крысы существование в ней "блобс" предполагал М. Наказава (1992).
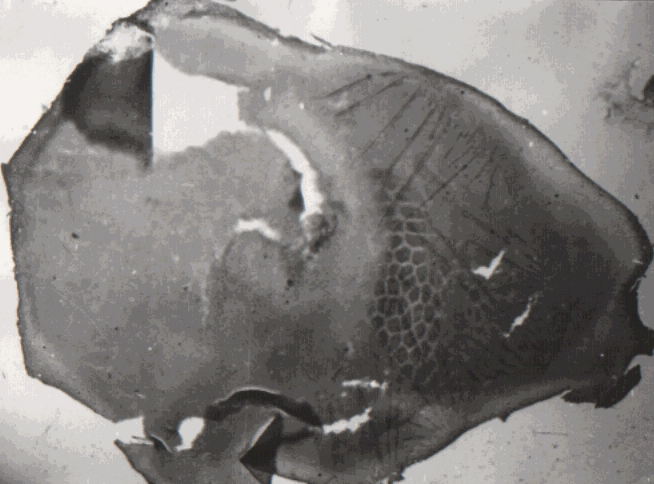
На рис. 2 демонстрируется фотография препарата, представляющая практически всю поверхность коры мозга белой крысы. Видно, что достаточно униформные группировки размером около 200мкм обнаруживаются по всему плащу коры. При этом надо учесть толщину среза - 75мкм, что естественно делает невозможным одинаковую резкость изображения в разных участках среза.
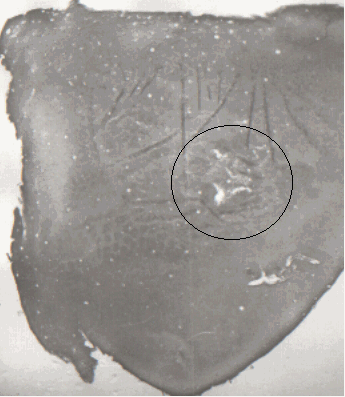
На Рис.3 - фотография представляющая "бочонки" соматосенсорной коры вместе с лежащими кпереди ассоциативными зонами - поля 5 и 7, лежащей рядом с "бочонковой" зоной соматосенсорной коры безбочонковой зоны, а ниже - слуховым полем.
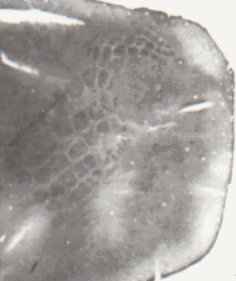
На Рис.4 иллюстрируется срез, включающий в себя участок моторной и - кпереди - премоторной коры мозга.
Видимо, не оставляет сомнений, что группировки нейронов с повышенной активностью цитохромоксидазы являются универсальным компонентом архитектоники по всему плащу новой коры. - Это для белой крысы. Мы однако упоминали, что такие же нейронные структуры нами были обнаружили прежде по всему плащу коры и в мозге морской свинки.
Отмеченные факты дают основания, чтобы рассматривать колонки коры с поперечником около 200 мкм, характеризующиеся повышенной активностью цитохромоксидазы, в качестве универсальных базовых модулей неокортекса.
Таким образом, с учетом упоминавшихся в преамбуле результатов исследования конструкции и механизмов функционирования базовых интегративных колонок можно заключить, что на примере коры мозга грызунов нам удалось выявить реальные универсальные базовые интегративные модули неокортекса.
Сделать это для мозга высших млекопитающих также на наш взгляд вполне реально (при условии правильного приготовления тангенциальных срезов), но видимо, из-за извилин и большой площади "плаща" неокортекса у высших млекопитающих, осуществить это будет возможно лишь по частям, путем сопоставления результатов различных исследований.
В заключение еще раз напомним, что в силу законов филогенеза - развитие всех млекопитающих от общего предка (Диамонд и Холл, Батуев и Куликов) и эмбриогенеза - развитие путем надстройки (принцип структурного дополнения - Богословская, Поляков), конструкция неокортекса на основе базовых универсальных интегративных модулей у любого млекопитающего должна иметь место с необходимостью и быть в принципе такой же, как это выявлено для грызунов в настоящей работе.
1. Hebb D. The organization of behavior. - N-Y, J. Wiley. - 1949.
2. Lorente de No R. The cerebral cortex: Architecture, intra-cortical connections and motor projection// Physiology of the Nervous System. - by J. F. Fulton, London: Oxford Univ. Press, 1938, Chapter 15.
3. Mountcastle V. B. Modality and topographic properties of single neurons of cat-s somatic sensory cortex// J. Neurophysiol., 1957, V.20, p.408-434.
4. White G. The formation of cell assemblies // Bull. Mathem. Biophysics - 1961. - V. 23. - P. 43-59
5. Шайбель М.Е., Шайбель А.Б. Структурный субстрат интеграции ретикулярной сердцевины ствола мозга// Ретикулярная формация мозга. - М.: Госмедиздат, 1962, С. 36.
6. Hubel D.H., Wiesel T.N. Receptive fields, binocular interaction and functional architecture in the cat-s visual cortex. - J. Physiol., V.160: 106-154 (1962).
7. Чораян О.Г. К функциональной нейроархитектонике // Физиол. журн. СССР, Т.54, N9, 1963, С.. 1026-1029.
8. Юнг Р. Интеграция в нейронах зрительной коры и ее значение для зрительной информации" // Теория связи в сенсорных системах. - М.: Мир, 1964, С.375.
9. Baumgartner G., Braun J.L/, Shulz A. Responses of single units of the cat visual system to rectangular stimulus patterns.// J. Neurophysiol., 1965, V.28, N 1, P.1-18.
10. Brown L. Shulz A. Responses of single units of the cat visual system to rectangular stimulus patterns// J. Neurophysiol., 1965, V. 28, N 1, p. 1-18.
11. Jung R. Neuronal Grundlagen des Hell-Dunkel Zone und der Farbwahrnehmung // Ber. Deutsche ophtal. Jes., 1965. B.66, s. 69-110.
12. Colonnier M.L. Structural design of the neocortex.// Brain and Consciouse Experience. N.Y., 1966. P.1-23.
13. Коган А.Б., Петунин Ю.И., Чораян О.Г. Исследование импульсной активности нейронов методами теории случайных процессов// Биофизика 1966, XI, N 5, с. 887-893.
14. Colonnier M., Rossignol S. Heterogenity of the cerebral cortex // Basic mechanisms of the epilepsies, Boston, 1969, p. 29-40.
15. Diamond I.T., Hall W.C. Evolution of neocortex.// Science, 1969, V.164, N 3877, P.251.
16. Mandle J. Localization of visual patterns by neurons in cerebral cortex of the cat// J. Neurophysiol., 1970, v. 33, N 6, p. 812-826.
17. Woolsy T.A. von der Loos H. The structural organization of layer 4 in the somatosensory region (SI) of mouse cerebral cortex.// Brain Res., 1970, V.17, N2, P.205.
18. Welker C. Microelectrode delination of fine grain somatotopic organization of SmI cerebral neocortex in albino rat// Brain Res, 1971, V. 26, N2, p. 255-275.
19. Подладчикова Л.Н. Динамика функциональной организации нейронов зрительной коры при изменении уровня ее возбудимостиь // Автореф. дисс. канд. биол. наук. - г.Ростов-на-Дону, 1972.
20. Чебкасов С.А. Пространственное распределение вызванных потенциалов и организация импульсных ответов нейронов зрительной коры мозга морской свинки. - Дисс. канд. биол. наук. - г. Ростов-на-Дону, РГУ, 1972. - 149с.
21. Подладчикова Л.Н., Кошуба Н.К. О пространственном распределении реакций нейронов стриарной коры морской свинки в зонах представительства центральной и периферической областей сетчатки. - Физиол. журн. СССР. - 1973. - Т. 59, N 3. - С.1183-1189.
22-Blakemore C. The Representation of visual space in area 17 of the cat. - Nova Acta Leopoldina. - 1973. - Band 37/2 . - N 208. - P. 155-171.
23. Тюрбеева М.Л. Гистологические показатели группирования нейронов в коре головного мозга кошки и морской свинки // Автореф. дис. канд. биол. наук. - Ростов-на-Дону: РГУ, 1973.
24. Батуев А.С., Бабминдра В.П. Некоторые морфофизиологические предпосылки межнейронной интеграции. // Системный анализ интегративной деятельности нейрона. - М.: Наука. 1974. С. 125-133.
25. Milner P. M. F. Model for visual shape recognition // Psychological Review. - 1974, Vol.81, N 6, P. 521-535.
26. Creutzfeldt O., Innocenti Y, Brook D. "Vertical organization in the visual cortex (area 17) in the cat", Exp. Brain. Res.- 1974, V. 21, N 3, p. 315-336.
27. Towe A. Notes of the hypothesis of columnar organization in somatosensory cerebral cortex // Brain. Behav. Evol., - 1975, V. 11, p. 16-47
28. Albus K.A. А quantative study of the projection area of the central and paracentral visual field in area 17 of the cat. The spatial organization of the orientation domain // Exp. Brain Res. - 1975, V. 24, N 2, p. 181-202.
29. Asanuma H. Resent developments in the study of the columnar arrangement of neurons within the motor cortex // Phisiol. Rev. - 1975, V. 55, N 2, P. 143-156.
30. Антонова А. М. Пространственная организация нейронных ансамблей слуховой коры мозга кошки.// Архив анат., гистол. и эмбриол., 1975. Т. LXVIII, N1, С. 73-78.
31. Чебкасов С.А. О некоторых закономерностях вариабельности первичных реакций нейронов зрительной коры мозга // В сб.: "Функционально-структурные основы системной деятельности и механизмы пластичности мозга": 1975. - вып. 4, С. 112-116.
32. Чебкасов С.А. О характере организации связей клеток нейронного ансамбля. - Ростов-на-Дону, НИИНК РГУ-каф.ФЧЖ, РГУ - I-я конференция-конкурс молодых ученых. - 1975. - Доклад - 9с.
33. Казаков В.П. Функциональные нейронные комплексы как основа интегративных процессов в ассоциативных областях коры головного мозга // Структурно-функциональные механизмы корковой интеграции. - Горький, 1976, - C. 102-104.
34. Creutzfedt O. The brain as a functional entity // Prog. Brain. Res. - 1976, V. 46, P. 451-462.
35. Creutzfeldt D. Afferent and intrinsic organization of the visual cortex columnar organization of continuous network in processing of information in the visual system // Ed. by Glezer V.D. - Leningrad, 1976, P. 223-224.
36. Антонова А.М. Структура неокортикальных клеточных ансамблей и принципы их пространственной организации // Структурно-функциональные механизмы корковой интеграции. - Горький: ин-т Мозга, Горьковский мед. ин-т, 1976. С. 10-14.
37. Сентаготаи Я, Арбиб М. Концептуальные модели нервной системы. - М.: Мир, 1976. - 198с.
38. Чебкасов С.А. О гетерогенности нейронного ансамбля и критериях его выявления. - Ростов-на-Дону. - Семинар НИИНК РГУ. - Доклад - 6с.
39. Чебкасов С.А., Тью-Тхи Хонг О закономерностях расположения нейронов зрительной коры с различной вариабельностью реакций. - Физиол. Журн. СССР - 1976, Т. 62, N 4, С. 468-470.
40. Lee B.B. et al. The depth distribution of optimal stimulus orientations for neuron in cat area 17. // Exp. Brain Res. - 1977, V. 27, N 3-4, P. 301-314.
41. Чебкасов С.А. К вопросу о пространственно-временной структуре первичной вызванной активности нейронов зрительной коры. - Физиол. журн. СССР - 1977. Т. 63, N 4. - С. 481-488.
42. Heggelund P., Albus K. Response variability and orientation discrimination of single cells in striate cortex of cat // Exp. Brain Res. - 1978, V. 32, N 2, P. 197-211.
43. Чебкасов С.А. О пространственно-временной мозаике нейронных ансамблей экранных структур мозга. - Ростов-на-Дону, НИИНК РГУ. - Доклад Ученому совету НИИНК, 1978 - 9с.
44. Szentagotai J. The neuron network of the cerebral cortex: a functional interpretation // Proc. Soc. London, 1978, V.201, N 1144, p.219-248.
45. Батуев А. С. Кортикальные механизмы интегративной деятельности мозга. - Л.:Наука, 1978.
46. Legendy C.R. Cortical columns and the tendency of neiboring neurons to act similarly/ - Brain Res. - 1978. - V. 158. - N 1. - P. 89-105.
47. Szentagothai J. The local neuronal apparatus of the cerebral cortex // Cerebral correlates of conscious experience. - Amsterdam, Elsevior North-Holland Biomed. Press, 1978, p.131-138
48. Сухов А.Г. К функциональной организации нейронов IV слоя соматосенсорной коры крысы // Структурно-функциональные основы организации мозга. - М.: Медицина, 1978: вып. 7, - С. 98-102.
49. Бондарь Г.Г., Чебкасов С.А., Подладчикова Л.Н. Пространственное распределение нейронов зрительной коры при стимуляции РФСМ. - Ростов-на-Дону: РГУ, 1979. Деп. ВИНИТИ, N 1532-79 В79.
50. Казаков В.Н., Шевченко Н.И., Пронькин В.Т. Колонки в коре головного мозга // Успехи физиол. наук, 1979, N 4. С. 96-115.
51. Антонова А.М. Нейронный ансамбль как полинейрональная системная единица // Интегративная деятельность нейрона. - М., 1979. - С. 76-78.
52. Антонова А.М. Нейронные ансамбли и проблема соматотопии.// Архив анат., гистол. и эмбриол., 1979, Т. 11. - С. 45-51.
53. Кузнецов С.А., Павлюк П.П., Цыбырнэ И.В. Корковая организация двигательнгой активности организма. - г. Кишенев, "ШТИИНЦА", 1979, - 142с.
54. Dreher B.A., Leventhal P.T., Hale P.T. Geniculate to cat visual cortex: a comparison of area 19 with 17 and 18. - J. Neurophphysiol. - 1980. - V. 44. - N 4. P. 804-826.
55. Чебкасов С.А. Некоторые принципы организации элементарных нейронных конструкций неокортекса. - Внутренний отчет о НИР в НИИНК РГУ. - 1980. - 175с.
56. Чебкасов С.А., Подладчикова Л.Н., Бондарь Г.Г. Элементарные ансамбли зрительной коры как локальные центры конвергенции различных информационных потоков // Материалы международн. конф. "Проблемы нейрокибернетики". - Ростов-на-Дону: РГУ, 1980, С. 48.
57. Белякова Г.В. Распределение нейронной активности в ЗКМ морской свинки при стимуляции специфических и каллозальных афферентов. - Дипломная работа, РГУ, кафедра ФЧЖ, 1980. - Руководитель - Л.Н. Подладчикова.
58. Кузнецов С. А. и др. Организация информационных процессов в корковом звене двигательного анализатора // Материалы международн. конф. "Проблемы нейрокибернетики". - Ростов-на-Дону: РГУ, 1980. - С. 29.
59. Ласков В.Н. О возможной организации системы кодирования местоположения
объектов в тактильном пространстве морды крысы // Материалы международн. конф. "Проблемы нейрокибернетики". - Ростов-на-Дону: РГУ, 1980. - С.30-31.
60. Wong-Riley M.T.T. & Welt C. Histochemical changes in cytochemical: changes in cytochrome oxidase of cortical barrels after vibriыsal removal in neonatal and adult mice // Proc. Natl. Acad. Sci. USA, 1980, V.77. N 4. P. 2333-2337.
61. Von der Malsburg C. Internal report in Max-Planck Institut fur Biophysikalische. - 1981.
62. Чебкасов С.А. Квантово-волновые представления об активности структурных колонок. Ансамбль-волна. - НИИНК РГУ, 1981. - Доклад городскому семинару по нейрокибернетике.
63. Богословская Л. С., Поляков Г. И. Пути морфологического прогресса нервных центров у высших позвоночных. ? M.: Наука, 1981. - 159с.
64. Бондарь Г.Г., Подладчикова Л.Н. Функциональная организация зрительной коры при стимуляции неспецифических структур среднего мозга и таламуса. - Ростов-на-Дону: РГУ, 1981. Деп. ВИНИТИ, N 1544-81 В81.
65. Чебкасов С.А. Метод микроэлектродного сканирования коры больших полушарий // Физиол. журн. СССР, 1981, N 12, С. 1886-1889.
66. Батуев А.С. Высшие интегративные системы мозга. - Л.: Наука, 1981. - 256с.
67. Эделмен Дж., Маунткасл В. Разумный мозг.- М.: Мир, 1981. - 135с.
68. Чебкасов С.А. Изучение пространственно-временной организации активности в нейронных ансамблях коры мозга. - Ростов-на-Дону: НИИНК, 1981. - Отчет о НИР по теме N ГР 81021765. - Этап 1981г. - 125с.
69. Abeles M. Role of the cortical neuron - integrator or coincidence detector? // Israel J. of Med. Science. - 1982. - V.18. - N 1. - P. 83-92.
70. Бабминдра В. П., Брагина Т.А. Структурные основы межнейронной интеграции. - Л.: Наука, 1982. - 163с.
71. Подладчикова Л.Н., Белякова Н.В., Бондарь Г.Г. О приуроченности афферентации из противоположного полушария к функциональным группировкам нейронов зрительной коры . - В сб.: "Взаимоотношения полушарий мозга". - Тбилиси, 1982. - С. 54-55.
72. Чебкасов С.А. Пространственное распределение гетерогенных афферентных влияний в зрительной коре мозга морских свинок // Журн. высш. нервн. деят., 1982, Т. 32, N 6, С.1156.
73. Чебкасов С.А. Об организации текто- и геникулокорковых афферентных влияний в связи с отдельными зонами формирования нейронных ансамблей// Вопросы эволюционной физиологии. - Л.: Наука, 1982, С. 360.
74. Батуев А.С., Куликов Г.А. Введение в физиологию сенсорных систем. - М.: Высш. Школа, 1983.
75. Szentagothai J., Erdi P. Outline of a general brain theory. - Budapest, Hungary, - 1983. - 101p.
76. Чебкасов С.А. Динамика пространственного распределения активности нейронов зрительной коры при стимуляции вспышкой диффузного света// Физиол. журн. СССР, 1983, Т.69, N 1, С. 46-54.
77. Чебкасов С.А. Влияние неспецифических ядер таламуса на отдельные локусы формирования ансамблей в зрительной коре мозга// Журн. высш. нервн. деят., 1983, Т.33, N 1, С.109-115.
78. Чебкасов С.А. Переработка информации о разных параметрах стимула в локусах формирования ансамблей // Материалы международн. конф. "Проблемы нейро-кибернетики". - Ростов-на-Дону: РГУ, 1983. ? С.60.
79. Чебкасов С.А. О возможном механизме интеграции нейронной активности в локусах формирования ансамблей. ? VII Всесоюзная с международным участием конференция по нейрокибернетике. - Ростов-на-Дону, 1983. - Круглый стол: "Формирование и свойства нейронных популяций". - Доклад. - 4с.
80. Чебкасов С.А. О пространственной организации нейронов зрительной коры мозга при стимуляции световыми пятнами // Физиол. журн. СССР. - 1983. - Т.69, - N8. - С.1099-1101.
81. Eccles J.C. The Cerebral Neocortex/ F Theory of Its Operation. - 1984. - 36p.
82. Goldman-Racick P.S. Modular organization of prefrontal cortex // Trennds in Neurosciences. - 1984. - V. 7, No 11, pp. 419-424.
83. Чебкасов С.А. О расположении и афферентных связях нейронов зрительной коры мозга, различающихся размерами рецептивных полей// Биол. науки. - 1984. - N3. - С.45-52.
84. Szentagotai J. Functional atnatomy of the visual cortex as cues for pattern recognition concepts // Pontif. Acad. Sci. Scr. Var., 1985, N 54, P. 39-52
85. Чебкасов С.А. Изучение свойств переработки информации в ансамблях нейронов и их мозаиках. - Ростов-на-Дону: НИИНК, 1985. - Отчет о НИР по теме N ГР 81021756. Этап 1985 г. - 26с.
86. Чебкасов С.А. О мозаичности расположения тормозных клеток зрительной коры // Нейронные механизмы коркового торможения. - Киев: Наукова Думка, 1985. - С.47.
87. Чебкасов С.А. О возможном дискретно-волновом механизме интеграции активности функционально различных нейронов // Зрение организмов и роботов. - Вильнюс, 1985. - С.76-77.
88. Чебкасов С.А. О расположении и афферентных связях нейронов зрительной коры мозга, различающихся размерами рецептивных полей // Биол. Науки, 1984. - N3. - С.45-52.
89. Chebkasov S.A. Influence of non-specific thalamic nuclei on separate ensembles formation loci in the visual brain cortex// Neurosci. A. Behav. Physiol. - 1984. - N3. - P.206-214.
90. Grossberg S. and Mingolla E. Neural dynamics of Form perception: boundary compeletion, illusory figers, and neon color spreading // Psychological Review 1985. Vol.92. .N2. 173-211.
91. Eccles J.C. The cerebral neocortex. - 1985. - V.2. - P.1-36.
92. Chebkasov S.A. The pattern of space distribution of visual cortex neuronal activity in stimulation with diffuse light flash // Neurosci. a. Behav. Physiol. - 1985. - N4. - P.295-306.
93. Чебкасов С.А. Изучение свойств переработки информации в ансамблях нейронов и их мозаиках. - Ростов-на-Дону, НИИНК РГУ. Отчет о НИР по теме N ГР 81021755. Этап 1985г. - 28с.
94. Стадников Е.Н. Способ резания коры мозга крысы в плоскости, параллельной мягкой оболочке. - Ростов-на-Дону, 1986. - Деп. ВИНИТИ N 6107-В86, 1986, 5с.
95. Rakic P. Intrinsic and extrinsic determinants of neocortical parcellations: a radial concept and model. - "Neuro-biol. Neocortexs Rept Dahlem. Workshop, Berlin, May 17-22, 1987". Chichester e.a. 1988. - P.5-27.
96. Rakic P. Specification of cerebral cortical Areas// Science. - 1988. - V.241, - N4862. - P.170-176.
97. Edelmen J.M. Neural Darvinism. - Basic book, Part two - The Extended Theory - P. 37-108. - N-E, 1988.
98. Walsh C. and Cepko C.L. Clonally zelated cortical cells show several migration patterns. - Science. - 1988. - V.241. - N 4871. - P. 1342-1345.
99. Kuljus R.O., Rakic P. - Soc. Neurosci. Abstr. - 1988. - V. 14.
100. Чебкасов С.А. О механизмах переработки информации в зрительной коре. - Ростов-на-Дону, 1988. - Деп. в ВИНИТИ, N 4548, В-88. - 38 с.
101. Чебкасов С.А. Об организации нейронных конструкций коры мозга - Ростов-на-Дону: РГУ, 1988. - Деп. в ВИНИТИ, N 2952, В-88. - 174с.
102. Никольс Дж. Динамика иерархических систем. Эволюционное представление. - М.: Мир, 1989. ? 488с.
103. Чебкасов С.А. Дипольная организация ориентационночувствительных модулей как пример структурной дополнительности нервных конструкций // Материалы международн. конф. "Проблемы нейрокибернетики". - Ростов-на-Дону: РГУ, 1989. - С. 60.
104. Чебкасов С.А. О назначении обратных