


Вопросы экологии применительно к лесному хозяйству
ЭКОЛОГИЯ
Контрольная работа № 1
Вопрос № 8.
Вода, как среда обитания животных организмов: плотность, давление, кислород, освещенность, солевой режим, течения, температура. Отличия от воздушной среды. Приспособления растений и животных.
Благодаря широкой распространённости воды и её роли в жизни людей, она издавна считалась первоисточником жизни. Представление философов античности о воде как о начале всех вещей нашло отражение в учении Аристотеля (4 в. до н. э.) о четырёх стихиях (огне, воздухе, земле и воде), причём вода считалась носителем холода и влажности. Вплоть до конца 18 века в науке существовало представление о воде, как об индивидуальном химическом элементе. В 1781—82 годах английский учёный Г. Кавендиш впервые синтезировал воду, взрывая электрической искрой смесь водорода и кислорода, а в 1783 году французский учёный А. Лавуазье повторив эти опыты, впервые сделал правильный вывод, что вода есть соединение водорода и кислорода. В 1785 году Лавуазье совместно с французским учёным Ж. Менье определил количественный состав воды. В 1800 году английские учёные У. Николсон и А. Карлейль разложили воду на элементы электрическим током. Таким образом, анализ и синтез воды показали сложность её состава и позволили установить для неё формулу H2O. Изучение физических свойств воды началось ещё до установления её состава в тесной связи с другими научно-техническими проблемами. В 1612 году итальянский учёный Г. Галилей обратил внимание на меньшую плотность льда сравнительно с жидкой водой как на причину плавучести льда. В 1665 году голландский учёный Х. Гюйгенс предложил принять температуру кипения и температуру плавления воды за опорные точки шкалы термометра. В 1772 году французский физик Делюк нашёл, что максимум плотности воды лежит при 4°С; при установлении в конце 18 века метрической системы мер и весов это наблюдение было использовано для определения единицы массы – килограмма. В связи с изобретением паровой машины французские учёные Д. Араго и П. Дюлонг (1830 г.) изучили зависимость давления насыщенного пара воды от температуры. В 1891—97 годах Д. И. Менделеев дал формулы зависимости плотности воды от температуры. В 1910 году американский учёный П. Бриджмен и немецкий учёный Г. Тамман обнаружили у льда при высоком давлении несколько полиморфных модификаций. В 1932 году американские учёные Э. Уошберн и Г. Юри открыли тяжёлую воду. Развитие физических методов исследования позволило существенно продвинуться в изучении структуры молекул воды, а также строения кристаллов льда. В последние десятилетия особое внимание учёных привлекает структура жидкой воды и водных растворов.
Физические свойства и строение воды. Важнейшие физические константы воды приведены в таблице 1. Тройная точка для воды, где находятся в равновесии жидкая вода, лёд и пар, лежит при температуре +0,01°С и давлении 6,03·10-3 атм.
Многие физические свойства воды обнаруживают существенные аномалии. Как известно, свойства однотипных химических соединений у элементов, находящихся в одной и той же группе периодической системы Менделеева, изменяются закономерно. В ряду водородных соединений элементов VI группы (H2Te, H2Se, H2S, H2O) температуры плавления и кипения закономерно уменьшаются лишь у первых трёх; для воды эти температуры аномально высоки. Плотность воды в интервале 100—4°С нормально возрастает, как и у огромного большинства других жидкостей. Однако, достигнув максимального значения 1,0000 г/см3 при +3,98°С, при дальнейшем охлаждении уменьшается, а при замерзании скачкообразно падает, тогда как почти у всех остальных веществ кристаллизация сопровождается увеличением плотности. Вода способна к значительному переохлаждению, т. е. может оставаться в жидком состоянии ниже температуры плавления (даже при —30°С). Удельная теплоёмкость, удельная теплота плавления и кипения воды аномально высоки по сравнению с другими веществами, причём удельная теплоёмкость воды минимальна при 40°С. Вязкость воды с ростом давления уменьшается, а не повышается, как следовало бы ожидать по аналогии с другими жидкостями. Сжимаемость воды крайне невелика, причём с ростом температуры уменьшается.
Таблица 1. Физические свойства воды
| Свойство | Значение Воздух для ср. |
Плотность, г/см3 | |
| лёд | 0,9168 (0°С) |
| жидкость | 0,99987 (0°С) 1,0000 (3,98°С) 0,99823 (20°С) 0,001248 (20°С) |
| Пар насыщенный | 0,5977 кг/м3 (100°С) |
| Температура плавления | 0°С |
| Температура кипения | 100°С |
| Критическая температура | 374,15°С —140,7°С |
| Критическое давление | 218,53 кгс/см2 3,7 Мн/м2 (37,2 am) |
| Критическая плотность | 0,325 г/см3 |
| Теплота плавления | 79,7 кал/г |
| Теплота испарения | 539 кал/г (100°С) |
Удельная теплопроводность, кал/(см·сек·град) | |
| лёд | 5,6·10-3 (0°С) |
| жидкость | 1,43·10-3 (0°С) 1,54·10-3 (45°С) |
| Пар насыщенный | 5,51·10-5 (100°С) |
Уд. электропроводность, ом--1·см-1 | |
| лёд | 0,4·10-8 (0°С) |
| жидкость | 1,47·10-8 (0°С) 4,41·10-8 (18°С) 18,9·10-8 (50°С) |
Удельная теплоёмкость Кал/(г·град) | |
| жидкость | 1,00 (15°С) 10,045·103 дж/(кг·К) |
| Пар насыщенный | 0,487 (100°С) |
| Диэлектрическая проницаемость | |
| лёд | 74,6 (°С) |
| жидкость | 81,0 (20°С) 1,000059 (0°С). |
| Пар насыщенный | 1,007 (145°С) |
| Вязкость, спз | |
| Жидкость | 1,7921 (0°С) 0,000171 (0°С) 0,284 (100°С) |
Поверхностное натяжение жидкой воды на границе с воздухом, дин/см | 74,64 (0°С) 62,61 (80°С) |
Показатель преломления (D — линия | 1,33299 (20°С) 1,00029 |
Примечание: 1 кал/(см·сек·град) = 418,68 вт/(м·К); 1 ом-–1·см-–1 = 100 сим/м;
1 кал/(г·град) =.4,186 кдж (кг·К); 1 спз = 10—3н·сек/м2; 1 дин/см = 10–3н/м.
Аномалии физических свойств воды связаны со структурой её молекулы и особенностями межмолекулярных взаимодействий в жидкой воде и льде. Три ядра в молекуле В. образуют равнобедренный треугольник с протонами в основании и кислородом в вершине (Рис. 1, а). Распределение электронной плотности в молекуле В. таково (Рис 1, б, в), что создаются 4 полюса зарядов: 2 положительных, связанных с атомами водорода, и 2 отрицательных, связанных с электронными облаками необобществлённых пар электронов атома кислорода. Указанные 4 полюса зарядов располагаются в вершинах тетраэдра (Рис 1, г). Благодаря этой полярности вода имеет высокий дипольный момент(1,86 D), а четыре полюса зарядов позволяют каждой молекуле воды образовать четыре водородные связи с соседними (такими же) молекулами (например, в кристаллах льда).м
Рис. 1. Структура молекулы воды: а — геометрия молекулы H2O (в парообразном состоянии); б — электронные орбиты в молекуле H2O; в — электронная формула молекулы H2O (видны необобществленные электронные пары); г — четыре полюса зарядов в молекуле H2O расположены в вершинах тетраэдра.
Вода входит в состав всех живых организмов, причём в целом в них содержится лишь вдвое меньше воды, чем во всех реках Земли. В живых организмах количество воды, за исключением семян и спор, колеблется между 60 и 99,7% по массе. По словам французского биолога Э. Дюбуа-Реймона, живой организм есть l'eau animée (одушевлённая вода). Все воды Земли постоянно взаимодействуют между собой, а также с атмосферой, литосферой и биосферой.
Вода в природных условиях всегда содержит растворённые соли, газы и органические вещества. Их количественный состав меняется в зависимости от происхождения воды и окружающих условий. При концентрации солей до 1 г/кг воду считают пресной, до 25 г/кг — солоноватой, свыше — солёной.
Наименее минерализованными водами являются атмосферные осадки (в среднем около 10—20 мг/кг), затем пресные озёра и реки (50—1000 мг/кг). Солёность океана колеблется около 35 г/кг; моря имеют меньшую минерализацию (Чёрное 17—22 г/кг; Балтийское 8—16 г/кг; Каспийское 11—13 г/кг). Минерализация подземных вод вблизи поверхности в условиях избыточного увлажнения составляет до 1 г/кг, в засушливых условиях до 100 г/кг, в глубинных артезианских вода минерализация колеблется в широких пределах. Максимальные концентрации солей наблюдаются в соляных озёрах (до 300 г/кг) и глубокозалегающих подземных водах. (до 600 г/кг).
В пресных водах обычно преобладают ионы HCO3-, Са2+ и Mg2+. По мере увеличения общей минерализации растет концентрация ионов SO42-, Cl-, Na+ и К+. В высо-коминерализованных водах преобладают ионы Cl- и Na+, реже Mg2+ и очень редко Ca2+. Прочие элементы содержатся в очень малых количествах, хотя почти все естественные элементы периодической системы найдены в природных водах.
Из растворённых газов в природных водах присутствуют азот, кислород, двуокись углерода, благородные газы, редко сероводород и углеводороды. Концентрация органических веществ невелика — в среднем в реках около 20 мг/л, в подземных водах ещё меньше, в океане около 4 мг/л. Исключение составляют воды болотные и нефтяных месторождений и воды, загрязнённые промышленными и бытовыми стоками, где количество их бывает выше. Качественный состав органических веществ чрезвычайно разнообразен и включает различные продукты жизнедеятельности организмов, населяющих воду, и соединения, образующиеся при распаде их остатков.
Первоисточниками солей природных вод являются вещества, образующиеся при химическом выветривании изверженных пород (Ca2+, Mg2+, Na+, К+ и др.), и вещества, выделявшиеся на протяжении всей истории Земли из её недр (CO2, SO2, HCI, NH3 и др.). От разнообразия состава этих веществ и условий, в которых происходило их взаимодействие с водой, зависит состав воды. Громадное значение для состава воды имеет и воздействие живых организмов.
В связи с существованием двух стабильных изотопов у водорода (1H и 2H, обычно обозначаемые Н и D) и трёх у кислорода (16O,17O и 18O) известно 9 изотопных разновидностей воды, которые находятся в природной воде в среднем в следующих соотношениях (в молярных %): 99,73 H216O; 0,04 H217O; 0,20 H218O, 0,03 HD’16O, а также 10-5—10-15%(суммарно) HD17O, HD18O, D216O, D217O, D218O. Особый интерес представляет тяжелая вода D2O, содержащая дейтерий. В водах Земли находится всего13—20 кг «сверхтяжёлой» воды. содержащей радиоактивный изотоп водорода — тритий (3H, или Т).
Вода в организме — основная среда (внутриклеточная и внеклеточная), в которой протекает обмен веществ у всех растений, животных и микроорганизмов, а также субстрат ряда химических ферментативных реакций. В процессе фотосинтеза вода вместе с углекислым газом вовлекается в образование органических веществ и, таким образом, служит материалом для создания живой материи на Земле.
Таблица 2. Содержание воды в различных организмах, их органах и тканях
| Организмы, органы, ткани | Содержание воды, % |
| Растения (наземные) | |
| верхушка растущего побега | 91—93 |
| листья . . . . . . . . . . . . . . . . . | 75—86 |
| Семена злаков . . . . . . . . . . . | 12—14 |
| Водоросли . . . . . . . . . . . . . . . | 90—98 |
| Мхи, лишайники . . . . . . . . . . | 5—7 |
| Медузы . . . . . . . . . . . . . . . . . | 95–98 |
| Дождевые черви . . . . . . . . . . | 84 |
| Насекомые | |
| взрослые . . . . . . . . . . . . . . . | 45—65 |
| личинки . . . . . . . . . . . . . . . . | 58—90 |
| Рыбы . . . . . . . . . . . . . . . . . . . | 70 |
Млекопитающие (в т. ч. человек) | 63—68 |
| скелет | 20—40 |
| мышцы | 75 |
| печень | 75 |
| Мозг человека | |
| серое вещество | 84 |
| белое вещество | 72 |
Вода обеспечивает тургор тканей, перенос питательных веществ и продуктов обмена (кровь, лимфа, сок растений), физическую терморегуляцию и другие процессы жизнедеятельности. Жизнь, вероятно, возникла в водной среде. В ходе эволюции различные водные животные и водные растения вышли на сушу и приспособились к наземному образу жизни; тем не менее и для них вода — важнейший компонент внешней среды. Жизнь без воды невозможна. При недостатке воды жизнедеятельность организмов нарушается. Лишь покоящиеся формы жизни — споры, семена — хорошо переносят длительное обезвоживание. Растения при отсутствии воды увядают и могут погибнуть, но чувствительность различных растений к недостатку воды неодинакова. Животные, если лишить их воды, быстро погибают: упитанная собака может прожить без пищи до 100 дней, а без воды — менее 10. Содержание воды в организмах велико (см. таблицу 2).
В жидкостях организма — межклеточных пространствах, лимфе, крови, пищеварительных соках, соке растений и др. — содержится свободная вода. В тканях животных и растений вода находится в связанном состоянии — она не вытекает при рассечении органа. Вода способна вызывать набухание коллоидов, связываться с белком и другими органическими соединениями, а также с ионами, входящими в состав клеток и тканей (гидратационная вода). Молекулы воды, находящиеся внутри клеток, но не входящие в состав гидратационных оболочек ионов и молекул, представляют иммобильную воду, легче гидратационной вовлекаемую в общий круговорот воды в организме.
Вода физиологически необходима любой протоплазме и с экологической точки зрения является лимитирующим фактором, как в наземных, так и в водных местообитаниях, если там ее количество подвержено резким изменениям (приливы, отливы) или происходит ее потеря организмом в сильно соленой воде осмотическим путем.
В наземно-воздушной среде этот абиотический фактор характеризуется величиной количества осадков, влажности, иссушающими свойствами воздуха и доступной площадью водного запаса.
Количество атмосферных осадков обусловлено физико-географическими условиями и неравномерно распределено на земном шаре. Но для организмов важнейшим лимитирующим фактором является распределение осадков по сезонам года. В умеренных широтах даже при достаточном количестве годовых осадков их неравномерное распределение может привести к гибели растений от засухи (лесных пожаров) или, наоборот, от переувлажнения. В тропической зоне организмам приходится переживать влажные и сухие сезоны, регулирующие их сезонную активность при постоянной почти круглый год температуре.
Адаптированные к условиям пустыни растения содержат ингибитор прорастания, который вымывается лишь при определенном количестве осадков, достаточном для вегетации (например, 10 мм) и тогда только прорастает. Начинается кратковременное «цветение пустыни» (обычно весной).
Иссушающее действие воздуха наиболее важное экологическое значение имеет для растений. Подавляющее большинство растений всасывает воду корневой системой из почвы. Иссушение почвы затрудняет всасывание. Адаптация растений к этим условиям – увеличение всасывающей силы и активной поверхности корней. Величина этой силы у корней умеренной зоны от 2 до 4 ∙ 106 Па, а у растений сухих областей – до 6 ∙ 106 Па. Как только выбрана вода в данном объеме, корни растут далее в глубь и стороны и корневая система может достигнуть, например, у злаков длины 13 км на 1000 см3 почвы (без корневых волосков).
Вода расходуется на фотосинтез, всего около 0,5 % всасывается клетками, а 97 – 99 % ее уходит на транспирацию – испарение через листья. При достатке воды и питательных веществ рост растений пропорционален транспирации, а ее эффективность будет наивысшей. Эффективность транспирации – это отношение прироста вещества (чистой продукции) к количеству транспирированной воды. Измеряется она в граммах сухого вещества на 1000 см3 воды. Для большинства растений она равна двум, т. е. на получение каждого грамма живого вещества тратится 500 г воды, даже для большинства засухоустойчивых. Основная форма адаптации – не снижение транспирации, а прекращение роста в период засухи.
В нижних ярусах тропических дождевых лесов, где 100%-ная относительная влажность, есть растения с приспособлениями для потери воды, а в пустынях у некоторых растений водный баланс не нарушается даже в период непродолжительной засухи, и т. д. В зависимости от способов адаптации растений к влажности выделяют несколько экологических групп, например: гигрофиты – наземные растения, живущие в очень влажных почвах и в условиях повышенной влажности (рис, папирус); мезофиты – переносят незначительную засуху (древесные растения различных климатических зон, травянистые растения дубрав, большинство культурных растений и др.); ксерофиты – растения сухих степей и пустынь, способные накапливать влагу в мясистых листьях и стеблях – суккуленты (алоэ, кактусы и др.), а также обладающие большой всасывающей силой корней и способные снижать транспирацию с узкими мелкими листьями – склерофиты.
Среди суккулентов наблюдается явление конвергенции – растения, относящиеся к разным видам, имеют практически одинаковую форму: африканский молочай и кактус имеют шарообразную форму, обеспечивающую наименьшую поверхность испарения.
Водная среда в среднем в 800 раз плотнее воздуха; этим объясняется возможность существования в воде животных, прозрачное, студенистое тело которых лишено прочных покровов или поддерживающего скелетного аппарата (медузы, сифонофоры, гребневики, сальпы и др.). Плотностью воды обусловлены и характерные для многих водных животных способы движения посредством ресничек или жгутиков (у большинства простейших, некоторых червей, кишечно-полостных и др., а также у личиночных форм губок, кишечно-полостных, червей, моллюсков, иглокожих и др.). Большая плотность воды позволяет очень мелким водным животным (планктон), способным лишь к слабым активным движениям, держаться в толще воды при помощи несложных приспособлений в виде крошечных пузырьков воздуха или капелек жира в их теле, или длинных, тонких выростов, увеличивающих поверхность тела. Только среди водных животных встречаются неподвижные прикреплённые формы, что обусловлено подвижностью воды и, следовательно, постоянным приносом находящейся в ней пищи в виде живых и отмерших планктонных организмов, так же как и разносом оплодотворённых яиц и личинок, который обеспечивает расселение прикрепленных форм.
Водные растения, также как и животные, обитающие в воде, в ходе эволюции своеобразно приспособились к данной среде обитания. Среди них различают гидрофиты — растения, погружённые в воду только нижней частью, и гидатофиты — растения, полностью или большей своей частью погруженные в воду. Обитание в водной среде обусловило особые черты организации водных растений: значительное увеличение поверхности тела в сравнении с его массой, что облегчает поглощение необходимых количеств кислорода и других газов, которых в воде содержится меньше, чем в воздухе. Увеличение поверхности тела достигается развитием больших тонких листьев (некоторые рдесты), расчленением листовой пластинки на тонкие нитевидные участки (уруть, роголистники, водные лютики); продырявливанием листьев или сильным развитием воздухоносных полостей и больших межклетников. У водных растений сильно развита разнолистность (гетерофилия): подводные, плавающие и воздушные листья на одном и том же растении значительно различаются как по внутреннему, так и по внешнему строению. Так, подводные листья не имеют устьиц; у плавающих на поверхности воды листьев устьица находятся только на верхней стороне, у воздушных листьев устьица — на обеих сторонах. Большая плотность водной среды обусловливает слабое развитие механических элементов в листьях и стеблях водных растений; немногочисленные механические элементы, имеющиеся в стеблях, расположены ближе к центру, что придаёт им большую гибкость. Так как интенсивность света в воде резко снижается, у многих водных растений в клетках эпидермиса имеются хлорофилловые зёрна. У водных растений слабо развиты или даже отсутствуют сосуды в проводящих пучках. Слабо развита и корневая система, а корневые волоски отсутствуют. Почти все водные растения — многолетники, размножающиеся вегетативно. Некоторые водные растения (наяда, роголистник) опыляются под водой; у других цветки поднимаются над водой, где и происходит опыление. Некоторые водные растения приспособились к периодическому высыханию водоёмов (например, частуха, стрелолист, жеруха).
Вопрос № 18
Динамика ценопопуляций растений: монокарпики и поликарпики, инвазия, запас диаспор в почве, семенная продуктивность, популяционные волны, самоизреживание, сукцессии, флюктуации, гомеостаз.
Ценопопуляция по определению – это составная часть биогеоценоза. Еще в XVII веке было установлено, что численность популяций растет по закону геометрической прогрессии, а уже в конце XVIII века Томас Мальтус (1766 – 1834 гг.) выдвинул свою известную теорию о росте народонаселения в геометрической прогрессии. Эта закономерность выражается кривой, изображенной на рис. 2.
На современном математическом языке эта кривая отражает экспоненциальный рост численности организмов и описывается уравнением:
Nt = N0 ert ,
где Nt – численность популяции в момент времени t;
N0 – численность популяции в начальный момент времени t 0;
e – основание натурального логарифма (2,7182);
r – показатель, характеризующий темп размножения особей в данной популяции.
Экспоненциальный рост возможен только тогда, когда r имеет постоянное численное значение, так как скорость роста популяции пропорциональна самой численности:
ΔN / Δ t = r N, где r = const.
Если численность отложить в логарифмическом масштабе, то кривая приобретает вид прямой линии (рис. 2, b).
Таким образом, экспоненциальный рост численности популяции – это рост численности ее особей в не изменяющихся условиях.
Условия, сохраняющиеся длительное время постоянными, невозможны в природе. Если бы это было не так, то, например, обычные бактерии могли бы дать такую массу органического вещества, которая могла покрыть весь земной шар слоем толщиной в 2 метра за 2 часа.
Однако такого в природе не происходит, так как существует множество ограничивающих факторов. Но есть примеры, когда при замедлении роста, т. е. при снижении r, экспоненциальный рост сохраняется, может он возникать и на коротких отрезках жизни популяций.
Чтобы иметь полную картину динамики численности популяции, а так же рассчитать скорость ее роста, необходимо знать величину так называемой чистой скорости воспроизводства (R0), которая показывает, во сколько раз увеличивается численность популяции за одно поколение, за время его жизни T.
R0 = Nr / N0 ,
где Nr – численность нового поколения;
N0 – численность особей предшествующего поколения;
R0 – чистая скорость воспроизводства, показывающая также, сколько вновь родившихся особей приходится на одну особь поколения родителей. Если R0 = 1, то популяция стационарная, численность ее сохраняется постоянной.
Скорость роста популяции обратно пропорциональна длительности жизни поколения:
r = ln R0 / T,
отсюда ясно, что чем раньше происходит размножение организмов, тем больше скорость роста популяции. Беря во внимание, например, этологическую структуру популяций монокарпических и поликарпических растений, выражающую характер их организации, можно заметить тот факт, что скорость роста популяции монокарпиков значительно ниже скорости роста популяции поликарпиков. Это объясняется тем, что
R0 м << R0 п ,
где R0 м – чистая скорость воспроизводства монокарпических растений;
R0 п – чистая скорость воспроизводства поликарпических растений.
Воздействие экологических факторов на скорость роста популяции может довести численность популяции до стабильной (r = 0), либо ее уменьшить. Например, к уменьшению численности популяции может привести воздействие инвазионных заболеваний, вызванных заражением растений паразитами животной природы. Инвазия может произойти активно, когда паразит нападает или внедряется в организм так называемого хозяина через поврежденные и неповрежденные ткани, или пассивно, когда паразит вносится в организм с водой, воздухом, почвенными растворами. Основными источниками заражения являются сточные воды предприятий пищевой и кожевенной промышленности, бытовые и промышленные свалки, кладбища, канализационная сеть, поля орошения и др. Из этих источников разнообразные органические соединения и патогенные микроорганизмы попадают в почву, горные породы и подземные воды. По данным санэпидстанций, патогенные кишечные палочки обнаруживаются в подземных водах на глубине до 300 метров от поверхности земли.
Состав и концентрация почвенного раствора определяют реакцию среды, показателем которой является величина pH. Наиболее благоприятной для растений и почвенных животных является нейтральная среда (pH = 7).
Однако в уменьшении численности популяций обычно играют роль вполне естественные ограничивающие или лимитирующие факторы (закон минимума Ю. Либиха, закон независимости факторов В. Р. Вильямса и закон толерантности В Шелфорда). В результате экспоненциальный рост замедляется или останавливается полностью и J – образная кривая экспоненциального роста как бы останавливается и выполаживается, превращаясь в так называемую S – образную кривую (рис. 3).
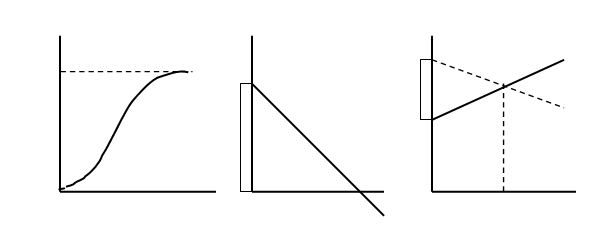
![]()
а r б b, d в
d
K
rmax
b
r max
N0 N N
0 Время, t 0 K 0 K
Рис. 3. Логистическая модель роста популяции: а – кривая роста численности (N); б – зависимость удельной скорости роста (r) от численности (N); в – зависимость рождаемости (b) и смертности (d) от численности; К – предельная численность.
В природе так и происходит: экспоненциальный рост наблюдается какое-то достаточно короткое время, после чего ограничивающие факторы его стабилизируют, и дальнейшее развитие популяции идет по логистической модели, что и описывается S – образной, или логистической кривой роста популяции.
В основе логистической модели (рис. 3) лежит простое допущение, что скорость роста популяции (ra) линейно снижается по мере роста численности вплоть до нуля при некой численности К. Итак, при начальной численности Na (близкой к нулю) скорость роста имеет максимальное значение rmax , а при N = K, r a = 0. В результате решения уравнения логистической кривой получается зависимость:
Nt = K / (1 + e α – r max ∙ t ),
где Nt – численность популяции в момент времени t;
α – постоянная интегрирования.
Величины К называют еще емкостью среды в отношении особей данной популяции. Здесь речь идет о биологической емкости среды – степени способности природного или природно-антропогенного окружения обеспечить нормальную жизнедеятельность (дыхание, питание, размножение, отдых и т.д.) определенному числу организмов и их сообществ без заметного нарушения самого окружения.
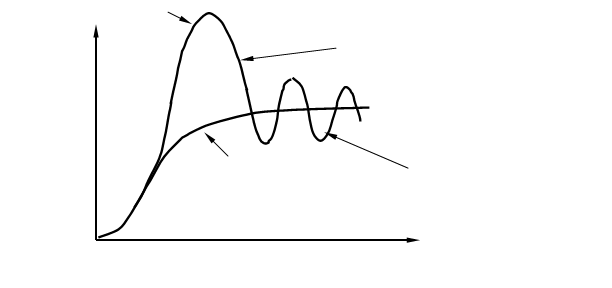
Однако плато на S – образной кривой далеко не всегда бывает гладким, потому что колебания численности происходят постоянно, что отражается в виде колебаний кривой вокруг асимптоты К (рис. 4), эти колебания называются популяционными волнами или флюктуациями численности. Данный термин был введён русским биологом С. С. Четвериковым в 1915 г. Подобные колебания численности могут быть сезонными или несезонными, повторяющимися через различные промежутки времени; обычно они тем длиннее, чем продолжительнее цикл развития организмов. Часто волны жизни сопровождаются колебаниями ареала популяций. Четвериков указал на эволюционное значение волн жизни в качестве фактора, могущего изменять направление и интенсивность отбора, а также концентрации генов, содержащихся в популяциях. Популяционные волны (один из 4 элементарных эволюционных факторов — мутационный процесс, популяционные волны, изоляция и естественный отбор). Основное значение популяционных волн сводится к случайным изменениям концентраций (особенно невысоких) различных мутаций и генотипов, содержащихся в популяциях, а также к ослаблению давления отбора при увеличении и его усилению при уменьшении численности особей в популяции. Под термином «популяционные волны» иногда (советский геолог Б. Л. Личков и некоторые другие ученые) также подразумевают этапы развития растительного и животного мира, примерно соответствующие смене геологических циклов.
Повышение популяцией
потенциальной емкости
«Крах» популяции
или вымирание
Число
особей
Потенциальная Популяционные
емкость волны
Время
Рис. 4. Преобразование J –образной кривой роста численности популяции в S – образную кривую при ограничивающем воздействии лимитирующих факторов.
Главенствующее значение для естественного возобновления ценопопуляции имеет запас диаспор в почве, наличие подроста и занос зачатков. В ненарушенной лесной среде практически исключена смена лесного типа растительности на луговой, степной или болотный. Если же ширина окна превышает две и более высоты сомкнутого окружающего полога, то возникает нелесная среда, где преимущество заселения получают вначале травяной и моховой типы растительности, и только потом заселяются пионерные береза, осина, иногда сосна, при этом частично или навсегда исчезают многие лесные виды растений, животных и, вероятно, микроорганизмов.
Можно наблюдать следующие закономерности возникновения возобновления сообществ. Почвенный запас семян хорошо защищен от поражения микроскопическими грибковыми организмами, неспорообразующими бактериями, спорообразующими бактериями, актиномицетами, но недостаточно – от представителей лесной фауны, особенно видов, питающихся в основном семенами. Случайным является попадание семян в оптимальные условия прорастания, совпадающие с генетической наследственностью. При прорастании семени проросток сразу вступает в тесные взаимоотношения с микробиоценозом, эдафотопом и климатопом, т.е. любое растение образует с биоценозом консорцию. В этот период наблюдается массовая гибель растений из-за несовпадения консорциативных и неблагоприятных экотопических связей. Объем пространства, занимаемый проростками и ювенильными древесными растениями, невелик, и конкуренции за свет и почву между ними нет. Ресурсы среды у них могут отнять мхи, травы и взрослые древесные особи. Это предопределяет всегда недостаточное число возобновления на начальном этапе. Даже количество возобновления, равное 2,5 млн. шт./га, означает 40 см2 (5 x 8 см), приходящихся на одно растение. Следовательно, энергию роста (бонитет) на начальном этапе полностью определяют почвенное богатство и сопряженность с микробоценозом. В это время закладывается жизненная стратегия роста и развития (так называемые классы Крафта – деление деревьев в лесу на пять классов по росту и развитию: прегосподствующие, господствующие, умеренно развитые, заглушенные и подпологовые) каждой особи. По мере роста особь увеличивает занимаемое ею пространство до начала смыкания кронами с другими особями, после чего начинается закономерная межвидовая и видовая ценопопуляционная конкуренция между особями в биогруппе вначале за свет надземного пространства, позже присоединяется конкуренция корневых систем особей за почвенные ресурсы. По отношению к каждой особи этот процесс получил название «давление среды», приводящее к изреживанию в биогруппах и в насаждении в целом. Выжившие особи увеличивают консорциативные связи,